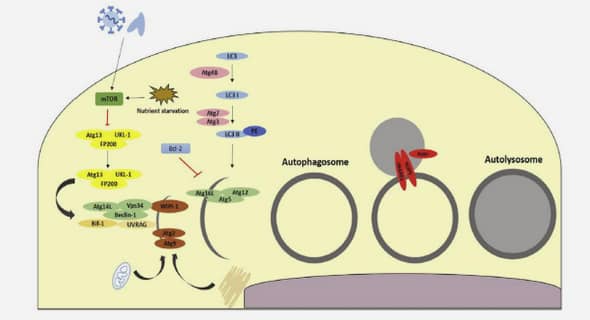
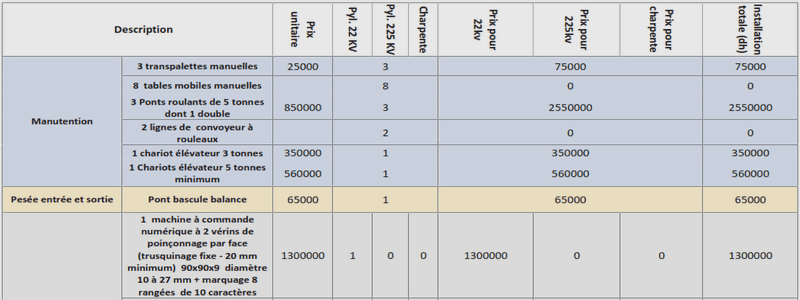
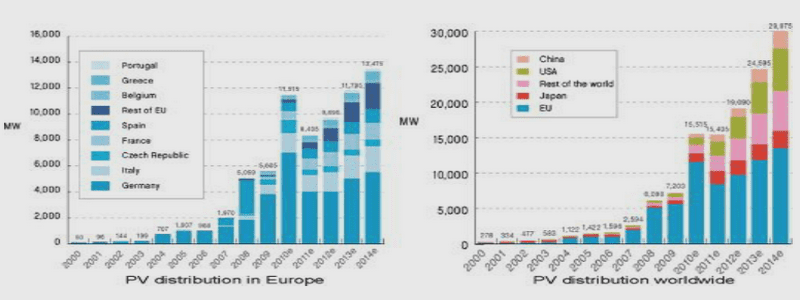
Etudes complémentaires par spectrométrie de masse
Les protéines Ku70p et Ku80p
Nous avons identifié la présence probable des protéines yKu70p (ou Ku70p) et yKu80p (ou Ku80p). Dans le cas de la protéine Ku70p, les résultats obtenus par spectrométrie de masse MALDI montrent un recouvrement par les peptides identifiés de 21.0 % de la séquence de cette protéine (figure 5.1.). Dans le cas de la protéine Ku80p, nous avons obtenu pour cette protéine un recouvrement de 20 % de la séquence (figure 5.2.). Indépendamment d’un taux de recouvrement important, trouver des éléments de séquence de ces deux protéines agissant habituellement de concert, accrédite l’hypothèse d’une association avec l’ADN Tlev. Jusqu’à présent, le complexe hétérogène Ku70p/Ku80p était connu pour s’associer au Chapitre V.Autres résultats – 101 – double brin et au brin G de l’ADN télomérique de Saccharomyces cerevisiae. Dans ces conditions, il n’est pas surprenant de retrouver ces protéines dans une association apparemment avec le brin C.Pour permettre de nous assurer de cette association avec la séquence Tlev, nous pourrions dépléter les extraits de levure en protéines Ku70p et Ku80p, par immunoprécipitation à l’aide d’anticorps spécifiques. Cette immunoprécipitation devrait, dans l’hypothèse d’une association de Tlev avec les protéines Ku70p et Ku80p, entraîner la disparition de l’une des bandes retardées d’intensité mineure observées après une migration électrophorétique non dénaturante.
La protéine Hsp60p
Nous obtenons une identification MALDI de la protéine Hsp60p, correspondant à un recouvrement de 37.8 % de la séquence. Nous en avons eu confirmation par identification par spectrométrie de masse en tandem d’un peptide spécifique de cette séquence (figure 5.3. en bleu). Cette protéine est localisée dans les mitochondries (Lithgow et al., 1995) où elle joue un rôle au niveau du repliement des protéines (Koll et al., 1992). Ses principales fonctions biochimiques connues sont celles de chaperone (Koll et al., 1992) et de protéine de choc thermique (Reading et al., 1989), mais à notre connaissance, aucune association de cette protéine avec de l’ADN n’a jusqu’à présent été mentionnée.
La protéine CBF3B
Nous obtenons une identification incomplète (11.3 % de séquence couverte) de la protéine CBF3B, sans confirmation par un autre moyen de séquençage. Cette protéine nucléaire fait partie du complexe protéique Cbf3 se liant à l’ADN centromérique (Lechner, 1994) et intervenant dans la mitose (Leopardi et al., 1996) et le contrôle du cycle cellulaire (Sorger et al., 1995).La répartition aléatoire des peptides recensés (figure 5.4.) et la masse native de cette protéine (71.349 Da) sont de bons indices de la présence de cette protéine dans les échantillons analysés. Nous n’avons pas pu déterminer la séquence d’un peptide par spectroscopie de masse en tandem et ces résultats sont insuffisants pour affirmer que cette protéine s’associe spécifiquement à la séquence Tlev. On sait cependant qu’elle se fixe à l’ADN centromérique du satellite CDEIII de Saccharomyces cerevisiae (Grady et al., 1992), qui est homologue au satellite III du centromère humain. La séquence de ce dernier est constitué de répétitions de CC (Fitzgerald-Hayes et al., 1982) et peut adopter une structure en motif i (Nonin-Lecomte & Leroy, 2001).
Purifications partielles et éléments d’identification
Dans le cadre de la recherche de protéines qui s’associeraient à la séquence télomérique de levure d[(CCCACA)3CCC], nous avons entrepris de développer un protocole de purification menant à l’isolement en conditions natives des protéines impliquées. Les difficultés rencontrées au cours de ces travaux sont en partie dues à l’instabilité constatée du complexe protéique en fonction de l’avancement des étapes de purification. 2-1- Eléments de purification Le travail présenté décrit les résultats obtenus lors de différentes phases de purifications partielles (pour chacune de ces étapes de purification, une étude préalable du comportement des protéines et de l’efficacité de la séparation a été conduite). 2-1-1- Chromatographie d’exclusion et chromatographie anionique Une première étape de séparation consiste à récupérer la fraction de chromatographie d’exclusion à 250 kDa (chapitre III-2-3-). La stabilité du complexe protéique est comparable à ce qu’elle était dans les extraits de protéines de levure (rappel : concentration en protéines = 1µg/µl). Le tampon d’élution est TBE 0.5x, pH 8 et NaCl 175 mM (utilisé pour neutraliser les interactions électrostatiques avec la résine. Le remplacement de NaCl par KCl a conduit à la perte de l’activité retardante.) . Nous observons un effet de sel de même genre lors de nos essais de séparation par chromatographie anionique : l’élution doit être réalisée en KCl, pour un pH compris entre 7.2 et 7.6. A l’opposé de ce qui se passe lors de la chromatographie d’exclusion, nous perdons l’activité retardée si nous utilisons du chlorure de sodium dans le tampon d’élution. Lors des expériences de chromatographie anionique, nous avons aussi déterminé que nous perdions l’activité retardée si la résine utilisée avait déjà préalablement servi. Après traitement dans des conditions d’élution dénaturantes (passage à pH 2 puis pH 10), nous observons le décrochage de Chapitre V.Autres résultats – 104 – produits que les conditions de chromatographie « natives » (gradient jusqu’à 2M KCl) ne permettent pas d’éluer. Ceux-ci pourraient être responsable de la perte de matériel constaté. Enfin, la chromatographie anionique consiste en une élution par pas de 50 et 100 mM de KCl plutôt que par gradient, afin d’obtenir une meilleure séparation entre les protéines présentes dans les extraits de levure. Dans ces conditions, on observe 90% du complexe protéique recherché dans la fraction correspondant au plateau à 300 mM KCL (figure 5.5.a.). Les protéines provenant de la fraction à 250 kDa de la chromatographie d’exclusion sont amenées aux conditions d’élution de la chromatographie anionique durant une étape de dilution – reconcentration (utilisation de tubes reconcentrateurs, chapitre II-10-) dans le tampon approprié (TBE 0.5x, pH 7.4, 50 mM KCl), sans perte de l’activité retardée. A la sortie de la colonne anionique, les fractions sont reconcentrées (volume 1 ml, volume final 10 µl) et dessalées avant d’en observer le contenu par migration électrophorétique native (figure 5.5.a) et par gel dénaturant de protéines (figure 5.5.b).
Développements ultérieurs
Devant l’instabilité du complexe protéique obtenu après ces deux étapes de purification ( 50% de perte du signal retardé après 2 h à 4°C et pas de bande retardée observée lors de migration électrophorétique native survenant soit après congélation à l’azote, soit après 1 nuit à 4°C), nous avons essayé d’identifier les protéines recherchées en profitant de la séparation complémentaire obtenue lors de la migration électrophorétique native. Pour cela, après une migration non dénaturante (réalisée à partir des fractions de la chromatographie anionique) le gel est séché sur une feuille de cellophane. Les bandes correspondant à l’activité retardée peuvent être découpées selon l’autoradiographie du gel, avant d’être insérées dans les puits pratiqués en haut d’un gel de protéines (figure 5.7.). Le découpage est tel que l’on retrouve 10% de l’activité retardée dans la piste de gauche du gel dénaturant, 85% dans la piste centrale et une fraction dans la piste de droite (estimations effectuées en mesurant la radioactivité au front de migration du gel de protéines). La bande de protéines de forte intensité (coloration saturée) observée dans la piste de gauche et la piste centrale correspond Chapitre V.Autres résultats – 107 – au doublet 9-10 de la figure 5.6. D’autres bandes sont également observées dans la piste centrale présentant une intensité relative beaucoup plus faible.