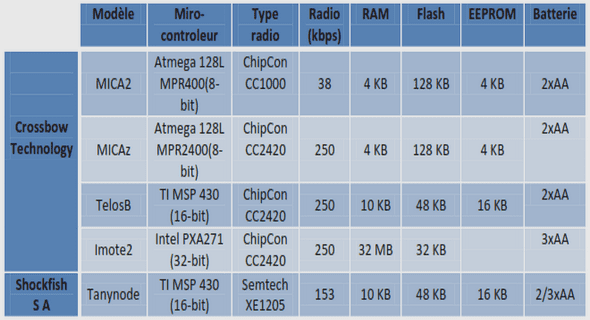
Les Treg Foxp3+
Caractérisation et rôle des Treg Foxp3+
Bien qu’il existe plusieurs types de Treg Foxp3+ comprenant les Treg CD4+, CD8+ ou encore possédant un TCRgd, nous ne détaillerons ici que les Treg CD4+ Foxp3+ qui sont les principauxreprésentants de cette classe de cellules régulatrices. Ces cellules, exprimant le facteur Foxp3+, sont des cellules caractérisées par des marqueurs clairs, même si chez la souris, leur identification est plus simple que chez l’homme. Ces cellules sont CD4+, CD45RBlow et expriment le CD25 qui est la chaine a du récepteur à l’interleukine-2. En association à la chaine b (CD122) et à la chaine g (CD132), le CD25 confère au Treg un récepteur avec une forte affinité alors même que le récepteur uniquement composé du CD25 ne possède qu’une faible affinité. Cette particularitéphénotypique leur permet d’être particulièrement sensibles à l’IL-2 même administrée à faible dose, ce qui ouvreun champ thérapeutique extrêmement vaste pour l’utilisation de cette cytokine en tant que thérapie immunorégulatrice (Klatzmann and Abbas, 2015).
Néanmoins, la dynamique des Treg sous IL-2 propre à chaque espèce ainsi que les variations individuelles en complexifient l’utilisation standardisée à des doses fixes(Churlaud et al., 2018) (Article n°2). Chez l’homme, à l’inverse des souris élevées en conditions exemptes de pathogènes spécifiques, on retrouve plus de LyT CD4+ exprimant le CD25 sans être pour autant des Treg, ce qui est très probablement la conséquence d’une exposition constante à des stimulus environnementaux. Néanmoins, le Foxp3 ainsi que l’absence d’expression du récepteur à l’interleukine-7 (CD127) restent des outils efficaces afin d’identifier et d’isoler les Treg chez l’homme. Les Treg Foxp3+ sont générés en premier lieu lors des phases de sélection clonale au sein du thymus et vont par la suite migrer en périphérie pour exercer leur fonction régulatrice. En dehors de ce mécanisme, les Treg FOPX3+ peuvent être également induits en périphérie à partirde LyT CD4+ naïfs par l’intermédiaire d’une faible exposition à un antigène ainsi que de la présence dans le microenvironnement d’IL-2 et de TGF-b.
Le développement de désordres immunitaires qu’il s’agisse de MAI ou d’allergie est fortement régulé par le rôle des Treg et plus particulièrement la balance entre les LyT effecteurs (Teff) et les Treg. En effet, on retrouve fréquemment une dérégulation de cette balance dans des MAI systémiques ou dans le diabète (Long et al., 2010). D’autre part, la restauration de l’équilibre Teff/Treg permet une amélioration clinique certaine par exemple dans l’arthrite ou l’allergie (Morgan et al., 2005; Yamashita et al., 2012). Demanière globale, ce déséquilibre peut être acquis soit par leur perte de fonction suppressive, soit par la diminution du nombre de Treg. C’est notamment le cas dans le syndrome IPEX avec l’absence d’expression de Foxp3 qui est essentiel pour le maintien de l’expression du CD25, du CTLA-4 ou encore de GITR (Hori et al., 2003). La délétion de ces cellules est également impliquée dans la perte de tolérance orale avec pour conséquence l’établissement d’une forte immunité de type Th2 et spécifique auxallergènes (Hadis et al., 2011). Concernant la perte de fonction suppressive, elle estretrouvée dans de nombreuses pathologies chez l’homme (e.g. DT1, SEP, PR, psoriasis).
Néanmoins sa mise en évidence reste compliquée car elle s’effectue exclusivement pardes cultures in vitro et ne constitue pas toujours le reflet de ce qui s’opère chez l’individu.
Mécanismes d’action des Treg Foxp3+
Les Treg Foxp3+ agissent par de multiples différences à la différence des autres populations de Treg (Figure 3). Cette polyvalence d’action leur permet d’agir à leur guise sur des effecteurs variés tout comme de modifier l’amplitude des réponses immunitairesà la base par leur action sur les DC.
Production de cytokines inhibitrices
Lorsque l’on regarde les Treg Foxp3+ thymiques, le rôle de l’IL-10 et du TGF-bdans leur efficacité suppressive semble mineur. En effet, dans des expériences in vitro, on observe une suppressionmaintenue même après déplétion de ces cytokines par utilisation d’anticorps spécifiques ou enutilisant des cellules incapables de produire ces cytokines (Dieckmann et al., 2001; Jonuleit et al., 2001). A l’inverse, des expériences in vivo en contexte pathologique, semblent démontrer que cesdeux cytokines restent indispensables dans l’activité des Treg (Hawrylowicz and O’Garra, 2005;Joetham et al.,2007). Ces observations effectuées dans le contexte de l’asthme allergiquedémontrent néanmoins que ce mécanisme ne peut se suffire pour induire une tolérance efficace. Eneffet, le transfert adoptif de Treg déficients pour l’IL-10 ne permet pas d’abroger l’effetprotecteur. En définitive, ces données supportent donc un rôle paracrine de l’IL-10 surles Treg Foxp3+ pour leur permettre de contrôler l’allergie et des recherches ultérieures ont démontré que ces Treg ne produisaient pas eux même cette cytokine à un niveausuffisant pour être suppresseurs (Rubtsov et al., 2008).
Le rôle du TGF-b demeure flou en partie à cause de sa présence sous deux formes : à la fois membranaire et soluble. Sous forme soluble, le TGF-b est impliqué à la fois dansle développement des Treg induits mais aussi dans le maintien de leur fonctionsuppressive. A l’inverse, sous forme membranaire, le TGF-b n’exerce pas ce rôle(Piccirillo et al.,2002) mais permet un contrôle direct des LyT CD8+ infiltrants lepancréas dans le diabète (Green et al., 2003). De plus, on retrouve le TGF-b à nouveau sous forme membranaire dans des exosomes en contexte tumoral avec un rôle dans l’accroissement des propriétés immunosuppressives des Treg. En définitive, cette molécule, quelle que soit sa forme, reste majeure dans l’action des Treg. En dernier lieu, l’IL-35 semble elle aussi jouer un rôle dans la prolifération et le maintien des fonctions suppressives des Treg (Collison et al., 2007). Cette cytokine est un membre de la famille de l’IL-12. Néanmoins, on remarque un manque important de recherches évaluant cette cytokine en conditions pathologiques et il n’est pas retrouvé d’action sur les Treg issus de l’Homme (Bardel et al., 2008).
Cytolyse
Loin d’être l’apanage seul des cellules cytotoxiques comme les NK ou les LyT CD8+ cytotoxiques (CTL), les Treg sont également décrits comme doués d’une capacitéde cytolyse avec la présence de granules de granzyme A, de granzyme B et de perforinequi peuvent être relargués suite à la fixation du CD18 (Grossman et al., 2004). Ces mécanismes de cytolyse sont mis en jeu notamment dans la suppression de l’activitécellulaire des LyB ainsi que des NK et CTL et en particulier dans un contexte tumoral (Vignali et al., 2008).
Privation en cytokines essentielles
Un des premiers rôle des Treg avancés après leur identification était de pouvoir capturer l’IL-2 grâce à leur très forte expression membranaire du CD25 (Thornton and Shevach, 1998). Cette observation précoce fut confirmée en 2007 (Pandiyan et al., 2007)où la privation en IL-2 dans le milieu est identifiée comme responsable de l’apoptose desLyT effecteurs (Teff), puis partiellement infirmée en avançant que ce mécanisme ne peutsuffire à lui seul (Oberle et al., 2007). En dehors de ce mécanisme historique, on reporte également l’implication d’une voie médiée par l’adénosine et les ectoenzymes CD39 et CD73. L’adénosine permettrait de supprimer les Teff à travers le récepteur adénosine 2A mais également d’induire plus de Treg par diminution de sécrétion d’IL-6 et hausse du TGF-b (Deaglio et al., 2007).
Action sur les cellules dendritiques
Les interactions entre les Treg et les DC sont au cœur des modulationsessentielles de l’activité immunitaire. Dans ce cadre, le CTLA-4 prend une part importante dans ce dialogue. Cette molécule fortement exprimée par les Treg permetune action primaire sur les DC par la liaison aux molécules de costimulation CD80 etCD86 en empêchant l’activation médiée par le CD28. Cette liaison est égalementresponsable de modifications enzymatiques au sein du microenvironnement de la DC.
De la même manière, LAG-3 joue un rôle sur les DC en bloquant leur maturation.
L’ensemble de ces moyens d’action sera détaillé plus précisément dans la description des points de contrôle immunitaires.
Les Treg sécréteurs d’interleukine-10 (Tr1)
Les cellules Tr1 sont décrites pour la première fois en 1997, dans les travaux de Maria Grazia Roncarolo, comme des LyT CD4+ capables de supprimer des réponses antigènes spécifiques et n’exprimant pas le Foxp3 (Groux et al., 1997). Ces cellules étaient à l’époque majoritairement caractérisées par leur sécrétion cytokinique propre, puis à lasuite d’un consensus international, par le phénotype CD4+ CD49b+ LAG-3+ CD226- (Abbas et al., 2013). Ces cellules restent néanmoins caractérisées par une productionimportante d’IL-10. En revanche, aucun facteur de transcriptionpropre ne peut êtreassocié à ces cellules malgré le fait que l’on retrouve une expression importante maisnon spécifique du répresseur de GATA3 (ROG) à la fois chez l’homme et chez la souris(Cobbold et al., 2003).
Fonctions des Tr1
A la différence des Treg Foxp3+, les Tr1 ne sont pas ou peu retrouvés au niveau systémique et leur présence est conditionnée par la présence d’un antigène spécifique, ce qui en fait des acteurs importants de la régulation immune locale (Zeng et al., 2015). De ce fait, l’étude des Tr1 s’est faite principalement en contexte pathologique et a permis d’identifier quatre mécanismes d’action principaux. Premièrement les Tr1 suppriment les Teff et modulent les DC par leur sécrétion d’IL-10. Sur les DC, l’IL-10 induitl’augmentation de l’expression de transcrits comme l’« Immunoglobulin-LikeTranscript » 3 et 4 (ILT3& ILT4) ainsi que la diminution d’expression des molécules decostimulationet de présentation antigénique. Les trois autres mécanismes retrouvéssont quant à eux partagés avec les Treg. Le deuxième est la production ainsi que lerelargage de granzyme B et de perforine lors de contacts avec les APC par l’intermédiaire entre autres du couple CD2 (LFA-2)/CD58 (LFA-3) (Magnani et al., 2011).Troisièmement, les Tr1 expriment les molécules CTLA-4, PD-1 et ICOS et peuvent doncmoduler l’activation des LyT naïfs par l’intermédiaire des DC (Gregori et al., 2012). En dernier, les Tr1 expriment les CD39 et CD73 et de ce fait modulent l’adénosine triphosphate extracellulaire et induisent un microenvironnement appauvri pour les Teff (Schuler et al., 2014). Cette propriété n’est cependant retrouvée que dans un contexte tumoral.
Induction de Tr1
Plusieurs techniques sont disponibles pour induire ces Tr1 dans une visée thérapeutique et pouvoir exploiter leur capacité suppressive ainsi que leur propension àmigrer au sein des tissus. Ces techniques sont majoritairement réalisées in vitro, car la génération de Tr1 in vivo semble extrêmement complexe à cause de la nécessité d’unemultitude de cytokines et de signaux d’activation d’intensité appropriée. L’étape clé dansla génération de ces cellules à partir de LyT CD4+ naïfs réside dans l’utilisation decertains sous-types de DC et particulièrement les iDC, les pDC ou les DC sécrétrices d’IL10. Dans cette optique et que ce soit chez l’homme ou chez la souris, il est possible d’utiliser des DC cultivées avec de la vitamine D3 et de la dexaméthasone (Barrat et al., 2002).Une seule étude fait état de la génération de Tr1 à partir de LyT CD4+ naïfs sansl’aide d’APC mais requiert l’utilisation de vecteurs viraux conférant la capacité de production de doses importantes d’IL-10 (Andolfi et al., 2012). L’étude qui parait la plus intéressante sur la génération deTr1 est celle publiée en 2016 qui évalue la génération de ces cellules in vivo par l’utilisation de nanoparticules vectorisant des complexes CMHII-peptides et de ce fait stimulant les cellules sans besoin de DC. Cette nanoparticuleinduit des cellules Tr1-like spécifiques de l’antigène, qui par la suite permettent de contrôler efficacement les manifestations auto-immunes dans différents modèles autoimmuns(encéphalomyélite expérimentale autoimmune (EAE), diabète, arthrite). Cette technique peut, par ailleurs, permettre de générer des Tr1-like spécifiques pour chaque antigène d’intérêt en créant les nanoparticules correspondantes (Clemente-Casares etal., 2016). En plus de contrôler les réponses cellulaires T, ces Tr1 sont également capables d’induire des LyB régulateurs (Breg) par l’intermédiaire de l’IL-10. Le rôle de ces Breg n’est pas clairement identifié mais reste cependant intéressant notamment dans des pathologies caractérisées par l’implication forte des immunoglobulines dans lapathologie. C’est notamment le cas dans l’allergie où ces Breg, par leur production d’IgGde type 4 contrôlent partiellement la réactivité aux allergènes.
Les Treg sécréteurs de TGF-b (Th3)
Les Treg polarisés vers un profil Th3 sont une population de cellules régulatrices très peu décrites en comparaison aux Treg Foxp3+ et aux Tr1. Ces cellules, identifiées pour la première fois au sein des ganglions mésentériques, sont impliquées dans la tolérance orale aux antigènes du soi (Chen et al., 1994). Par la suite, une meilleure caractérisationde ces cellules sera possible grâce aux travaux de recherche soulignant la dépendance au TGF-b pour leur action régulatrice et à l’expression membranaire en association avec le« latency-associated peptid » ou LAP à la surface de ces Th3 (Oida et al., 2003). Grâce à cette identification phénotypique claire (CD4+ CD25-LAP+), les Th3 sont étudiés dans plusieurs modèles de pathologies auto-immunes comme la colite ou l’EAE, qui est le parallèle murin de la SEP. De manière intéressante, ces cellules sont retrouvées chez l’Homme à la fois en contexte physiologique (Gandhi et al., 2010) mais également en contexte tumoral (Scurr et al., 2014). Si en situation classique, ces cellules permettent uncontrôle fin des réponses immunitaires, en contexte tumoral les Th3 sont clairement délétères avec même un rôle suppresseur supposé supérieur à celui des Treg. Il restecependant une grande inconnue dans le mécanisme d’action des Th3, dans la mesure oùla relation entre leur effet suppresseur et les Treg Foxp3+ n’est pas renseignée. En effet,le rôle majeur du TGF-b reste celui d’induire des Treg Foxp3+ à partir de LyT CD4+ naïfs.De plus, l’absence de facteurs de transcription identifiés pour les Th3, à la différence desTh1, Th2 et Treg avec respectivement T-bet, GATA3 ou Foxp3, complexifie la caractérisation complète de ces cellules. C’est donc probablement pour l’ensemble de ces raisons, que les développements de vaccination tolérogénique détaillés par la suite se concentrent principalement sur l’induction de Treg Foxp3+ et Tr1.
Pour clore cette description des acteurs clés de la vaccination tolérogénique, il estimportant de rappeler les deux principaux maitres d’œuvre de la tolérance immunitaireque sont les DC et les Treg. Bien qu’il reste encore de nombreuses voies d’action encore non décrites, il est aisé de se rendre compte du rôle primordial des molécules decostimulation, des cytokines et autres molécules régulatrices modulées par les cellules dendritiques. De leur côté, les Treg s’appuient sur des mécanismes divers, à la fois par contact et à distance des cellules effectrices. Ce système complexe représente une voie d’action thérapeutique majeure pour la vaccination tolérogène. En effet, la majorité des développements spécifiques d’un antigène vont s’atteler à moduler les DC avec en ligne de mire une action sur les Treg.