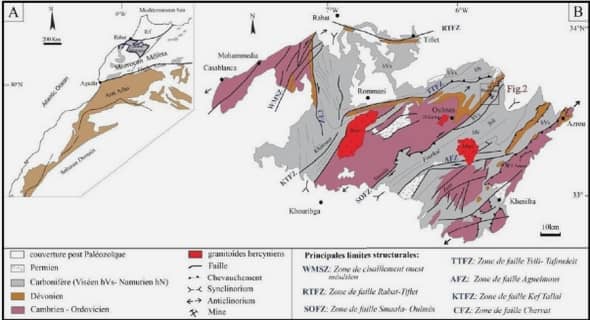
Méthodologie de détection de QTL par analyse de liaison revue bibliographique
Le génome correspond à l’intégralité du matériel génétique d’un individu, organisé en chromosomes. Les chromosomes, molécules d’Acide DésoxyriboNucléique (ADN), sont associés chez les espèces diploïdes et à reproduction sexuée, par paires de chromosomes homologues (l’un provenant du père et l’autre de la mère). En absence d’anomalie, on compte 40 paires chez les canards, 32 chez les chevaux, 30 chez les bovins et caprins, 27 chez les ovins et 23 paires chez l’Homme (encyclopédie universalis). Les chromosomes sont composés de séquences codantes (transcrites en ARN messager puis traduites en protéines) et de séquences non-codantes (non transcrites ou non traduites). Le gène appartient aux séquences codantes : c’est une unité d’information génétique transmissible, codée par une succession de bases nucléotidiques ayant 4 formes possibles (adénine, guanine, cytosine et thymine). La région du chromosome où se situe un gène particulier est appelée son locus. Chaque gène possède différentes versions appelées allèles. Si tous les individus d’une même espèce ont les mêmes gènes, chacun a un ensemble spécifique d’allèles. Un individu donné possède, pour chaque gène, un allèle reçu de son père et un allèle reçu de sa mère : c’est son génotype au locus. La séquence ordonnée des allèles de plusieurs locus adjacents forme un haplotype. Chez les espèces diploïdes, chaque individu a deux séries d’haplotypes, l’une héritée de son père et l’autre héritée de sa mère. L’analyse de liaison visant à mettre en évidence les portions de génome (QTL pour Quantitatif Trait Loci ou locus à caractère quantitatif) impliquées dans le déterminisme des caractères d’intérêt a été rendu possible par le développement des cartes génétiques de marqueurs. Il s’agit d’identifier les locus dont la variation allélique est associée à une variation d’un caractère d’intérêt.
Cette détection de QTL nécessite la mise en place de dispositifs animaux spécifiques. Deux types de populations peuvent être utilisés : les populations « inbred », lignées consanguines aux allèles fixés, et les populations « outbred » chez lesquelles les génotypes sont variés. Ces dispositifs font intervenir plusieurs générations d’individus (aux moins deux) composés de plusieurs familles de pères et/ou mères. La cartographie de QTL est d’autant plus précise qu’il y a de parents informatifs, c’est-à-dire hétérozygotes aux marqueurs et aux éventuels QTLs associés. Les croisements entre lignées « inbred » sont de ce point de vue très efficaces. Puis il est nécessaire de réaliser i) le marquage moléculaire de la génération des descendants et, quand c’est possible et utile, de générations antérieures. Il s’agit du génotypage des individus pour un réseau de marqueurs génétiques polymorphes régulièrement espacés sur le génome et repérables dans une carte génétique, et ii) le phénotypage des individus de la dernière génération c’est-à-dire la mesure des caractères d’intérêt. aisément détectable, située dans un locus unique et qui se transmet selon les lois mendéliennes. Il est utilisé pour « baliser » le génome. Comme cela sera décrit au paragraphe 3, l’ordonnancement de ces marqueurs et l’estimation de leurs distances génétiques aboutissent à la construction des cartes génétiques. Les marqueurs génétiques permettent de suivre la transmission d’un segment de chromosome d’une génération à l’autre. En effet, si un allèle X porté par un individu existe chez son père mais pas chez sa mère, l’individu l’a reçu de son père.
La probabilité pour que deux loci différents situés sur un même chromosome soient séparés par une recombinaison réciproque (crossing-over) entre ce chromosome et son homologue au cours de la méiose, est d’autant plus grande que ces loci sont plus éloignés l’un de l’autre. Si les loci sont proches ou s’il y a un nombre pair de recombinaisons entre eux, ils restent groupés sur le même chromosome dans le gamète produit par cette méiose. De tels loci, qui tendent à être hérités ensemble, sont dits liés. S’il y a un nombre impair de crossing-over entre eux, une recombinaison est observable. La proportion de recombinaison entre deux locus est une mesure de leur éloignement. Des gènes sont considérés comme liés si la probabilité de recombinaison est inférieure à 0,5. Pour des gènes non liés, la probabilité de recombinaison est égale à 0,5. Cependant, cette mesure ne possède pas les propriétés mathématiques des distances, mais, l’espérance du nombre de recombinaisons possède ces attributs. La construction d’une carte génétique se fait en deux grandes étapes. On constitue d’abord des groupes de liaison, c’est-à-dire des ensembles de marqueurs liés entre eux. Pour qu’un marqueur donné soit affecté à un groupe de liaison, il doit être lié à au moins un des marqueurs du groupe. Un groupe de liaisons correspond à un chromosome ou à un morceau de chromosome quand les marqueurs qui le composent sont, au moins en partie, ancrés sur ledit chromosome. Pour constituer ces groupes, on utilise la probabilité de recombinaison (r) entre les marqueurs deux à deux, en testant, grâce aux méthodes de maximum de vraisemblance, l’hypothèse H0: r = 0,5 c’est-à-dire qu’« il y a indépendance des marqueurs » contre l’hypothèse H1: 0 ≤ r < 0,5 c’est-à-dire que « les deux marqueurs sont liés ». Ensuite les marqueurs sont ordonnés à l’intérieur de chaque groupe de liaison, c’est-à-dire qu’on cherche l’ordre le plus vraisemblable de ces marqueurs.