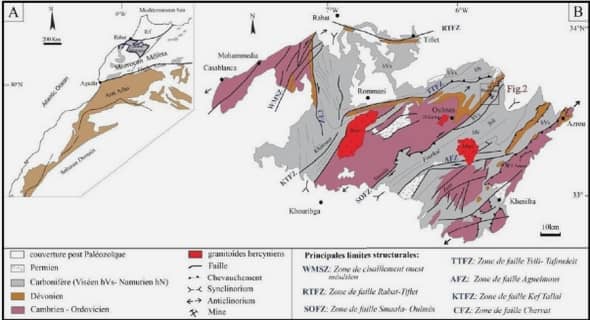
Une exploration multidimensionnelle des phénomènes expérientiels mnésiques
Les effets à l’échelle neuronale de la stimulation
Les effets neuronaux de la microstimulation
Dans les années 60 et 70, des expérimentations sur des coupes de cerveau animal ont pu démontrer que le premier effet de la stimulation électrique dans le système nerveux central était l’induction (génération) de potentiels d’action au niveau des axones (Ranck, 1975). Quand une stimulation extracelullaire est appliquée en substance grise, ce sont d’abord les axones qui sont excités plutôt que les corps cellulaires (Nowak & Bullier, 1998b, 1998a). A ce jour, plusieurs résultats suggèrent donc que le site d’activation directe des neurones est le segment initial de l’axone et les nœuds de Ranvier, qui ont les plus hauts taux de concentration en canaux sodiques et donc le plus haut niveau d’excitabilité. La contribution des dendrites et des corps neuronaux n’est que mineure comparativement aux axones dans l’activation neuronale (Ranck, 1975; Rattay, 1999; Rattay & Wenger, 2010). Le principe fondamental de l’application d’un courant électrique sur un processus neuronal est d’initier une réponse fonctionnelle en dépolarisant la membrane de cellules excitables. Cette 78 dépolarisation est produite par un flux de courant ionique entre deux électrodes (ou plus), dont l’une est très proche du tissu cible. L’application du courant électrique induit un changement de voltage transmembranaire qui entraine l’ouverture des canaux sodiques voltages dépendants sur un axone. Une stimulation électrique externe peut générer artificiellement des potentiels d’action sur une fibre nerveuse : les courants cathodiques ou anodiques ont pour répercussion une redistribution des charges le long de la membrane. La stimulation cathodique entraine une accumulation des charges à l’extérieur de la membrane et donc dépolarise la membrane sous l’extrémité de l’électrode. Une hyperpolarisation se produit à une certaine distance du site de stimulation par voie de conséquence, en raison d’un mouvement de charge positive vers l’espace intracellulaire. Une stimulation anodique produit l’effet inverse (une hyperpolarisation sous la membrane et une dépolarisation à distance) (Vincent et al., 2015). Les SEIC bipolaires, les plus couramment employées, sont utilisées pour obliger le courant à ne se propager que dans une seule direction. La fibre nerveuse reçoit alors des stimulations cathodiques et anodiques. Une extrémité de l’électrode délivre un courant négatif, la cathode, et l’autre un courant positif (l’anode). En augmentant la largeur de l’hyperpolarisation, c’est-à-dire l’amplitude du courant, le potentiel d’action qui est généré par la cathode peut être bloqué (Ranck, 1975; Vincent et al., 2015). Chez l’Homme, ces SEIC bipolaires sont délivrées entre deux contacts de la macro- ou micro-électrode. La phase cathodique du pulse de stimulation génère une dépolarisation membranaire dans les nœuds de Ranvier à proximité du contact de l’électrode. La magnitude de la dépolarisation est alors dépendante de la forme et de la trajectoire de l’axone, ainsi que de sa proximité avec l’électrode de contact. Si l’amplitude de stimulation est suffisante pour générer une dépolarisation qui dépasse le seuil d’activation de l’axone, un potentiel d’action sera généré et pourra se propager dans les deux directions à la fois orthodromique et antidromique, avec une efficacité dans ces deux directions (McIntyre & Anderson, 2016). Une certaine quantité de courant est nécessaire pour moduler/activer directement un neurone : elle est proportionnelle au carré de la distance entre le neurone (son corps cellulaire) et l’extrémité de l’électrode. Cette relation a été obtenue par des études chez le rongeur, le chat ou les primates sur des neurones corticaux ou cortico-spinaux en microstimulation (Stoney et al., 1968 ; Asanuma et al., 1976). Une fois que les potentiels d’action ont été générés, leur propagation vers la terminaison synaptique de l’axone entraine la libération synaptique de neurotransmetteurs. Parce que les éléments axonaux intrinsèques sont abondants dans le néocortex et la plupart des autres structures de la substance grise du système nerveux central, on doit s’attendre à ce que les effets antidromiques 79 locaux et/ou les effets orthodromiques synaptiques exercent la majeure partie de l’effet de stimulation même si seulement un petit nombre de corps neuronaux ont été stimulés directement. La stimulation électrique doit également être suffisamment longue et intense pour induire un changement du potentiel membranaire qui dépasse le seuil naturel d’excitation neuronale, selon une loi du tout ou rien. L’intensité de stimulation nécessaire diminue si la durée de stimulation augmente (Vincent et al., 2015). Il a longtemps été retenu comme règle générale que les microstimulations électriques activaient une sphère de neurones autour de l’extrémité de l’électrode, cette sphère augmentant en taille avec l’augmentation du courant (Stoney et al., 1968; Tehovnik, 1996). Selon cette hypothèse, on estime la population de neurones directement activée par une petite impulsion de courant (par exemple, entre 10 et 100 µA et pulse de 0,2 ms) se trouve dans un rayon autour de l’électrode de 100 à 450 micromètres (Stoney et al., 1968) et qu’elle excite ainsi quelques milliers de cellules directement (Tehovnik, 1996) Des données plus récentes, utilisant la microstimulation et l’imagerie calcique, suggèrent toutefois que des SEIC à faible intensité entrainent l’activation d’une population de neurones plus éparpillée et distribuée (Histed et al., 2009). Les effets de la SEIC seraient très sensibles à la région cérébrale qui est stimulée sur une échelle spatiale très limitée. Par exemple, chez des singes préalablement entrainés à une tâche de mémoire de reconnaissance visuelle avec présentation d’objets, la microstimulation électrique d’un « hotspot » bien précis du cortex périrhinal où se trouvent un grand nombre de neurones spécifiques d’un stimulus, permet aux singes de reconnaitre l’item comme familier. Mais stimuler juste à la périphérie de ce même hotspot (donc avec une variation de la position de l’électrode de stimulation à l’échelle micrométrique) entraine l’effet inverse, en leur faisant reconnaitre à tort l’item (pourtant préalablement présenté) comme nouveau (Tamura et al., 2017). Les auteurs font l’hypothèse que, sur la base des nombreuses connexions inhibitrices intrinsèques et locales des neurones du hotspot avec des neurones voisins au sein même du cortex périrhinal et des projections à plus longue distance vers des régions adjacentes, la stimulation électrique de ce hotspot pourrait inhiber les régions voisines en périphérie ce qui n’aurait pas d’impact sur les performances à la tâche. Tandis que stimuler les neurones en périphérie viendrait inhiber ceux du hostpot spécifique du stimulus avec pour conséquence l’induction d’un biais. Il faut enfin ajouter un effet potentiel de la SEIC sur des cibles non neuronales. A titre d’exemple, la glie est très probablement directement et indirectement influencée par la stimulation. De plus en plus de preuves suggèrent un rôle important du tissu non neuronal dans la modulation des cascades moléculaires et donc de l’activité neuronale, induite par la stimulation électrique cérébrale (McIntyre & Anderson, 2016). 80 Dans tous les cas, chaque potentiel d’action peut entrainer des centaines d’événements synaptiques en raison de l’importante arborisation axonale propre à chaque neurone.
Les effets neuronaux de la stimulation avec macroélectrodes
Ces résultats obtenus par l’étude de la microstimulation sont difficilement généralisables à toutes les structures cérébrales, puisque la densité et la composition neuronale varient en fonction des structures cérébrales. Plus largement la généralisation à la macrostimulation de ces observations à l’échelle neuronale reste encore acrobatique et hypothétique. S’il est possible d’obtenir une estimation correcte de l’extension du courant efficace et de la charge de courant délivré à l’extrémité d’une microélectrode (Tehovnik et al., 2006) ou entre deux macrocontacts (Tableau – Annexe 1), un certain nombre de questions sur l’étendue et le mode de propagation du signal et de ses effets après une SEIC utilisant des macroélectrodes ne sont pas résolues. Est-ce qu’il reste au niveau local, entre ces deux macrocontacts ? Est-ce qu’il existe un effet sur des régions éloignées ? Par l’étude de phénomènes lumineux induits par la SEIC du cortex visuel en électrocorticographie et une corrélation avec la position rétinotopique de ces stimulations dans les champs récepteurs, Winawer et Parvizi ont démontré qu’il existait une relation entre la magnitude de la charge de courant délivrée par la macrostimulation et la taille de l’aire corticale affectée (Winawer & Parvizi, 2016). On pourrait donc penser, en se limitant aux conclusions de cette étude en électrocorticographie, à une simple propagation en tache d’huile de la masse neuronale activée parallèlement à l’importance de la stimulation électrique. Mais ces données ne sont pas totalement en correspondance avec celles obtenues lors de la microstimulation. L’utilisation de l’imagerie calcique (two-photon calcium) chez l’animal après microstimulation (une méthode permettant d’observer l’activation des neurones sans être gêné par les artéfacts électriques de saturation du signal au site de stimulation) a démontré que le principal effet d’une augmentation de la charge du courant serait plutôt une augmentation de la densité de neurones au sein d’une région qui est définie par les projections axonales, donc éparses et non limitées à un simple périmètre autour de l’électrode. Les effets se propagent bien à distance de l’électrode (Histed et al., 2009). Cette distance peut être importante, puisque la microstimulation aurait la capacité d’inhiber des régions corticales dont on aurait stimuler les afférences (Logothetis et al., 2010). 81 Logothetis et al. (2010) ont employé simultanément la microstimulation électrique, l’électrophysiologie et l’IRM fonctionnelle pour démontrer que la stimulation pouvait engendrer une activation comme une inhibition de l’activité dans des structures cérébrales dont les afférences ont été stimulées, dépendant du site de stimulation et de la fréquence de stimulation. Dans l’exemple spécifique de la SEIC via des macroélectrodes du noyau sous-thalamique (NST) dans la maladie de Parkinson, en prenant en compte entre autres la densité neuronale du noyau sous-thalamique et du globus pallidal interne (GPI), ainsi que le nombre de synapses du NST au GPI estime qu’un pulse de SCP pourrait théoriquement entrainer 60 millions d’événements synaptiques dans le globus pallidum par stimulation directe du NST (McIntyre & Anderson, 2016). En résumé, en restant prudent car certains résultats ont été obtenus en microstimulation (Histed et al., 2009; Logothetis et al., 2010) et d’autres par estimation et simulation (McIntyre et al., 2004) , les données actuellement disponibles suggèrent qu’une augmentation de la charge de courant délivrée entraine non seulement une augmentation du nombre de neurones activés ou plutôt modulés au sein de la région stimulée mais également une extension de cette modulation au travers de connections fonctionnelles dans des régions éloignées. Ce n’est que l’année dernière qu’une étude chez des patients épileptiques pharmacorésistants a réussi à coupler l’IRM fonctionnelle à des macrostimulations en électrocorticographie , démontrant des modifications de connectivités à large échelle de SEIC dont l’intensité n’était pourtant pas suffisante pour induire des effets cliniques et comportementaux visibles (Oya et al., 2017).
Abréviations |