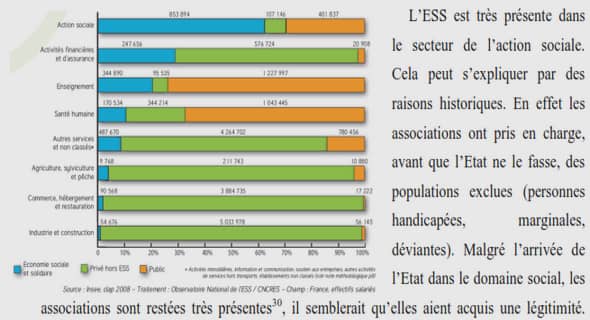
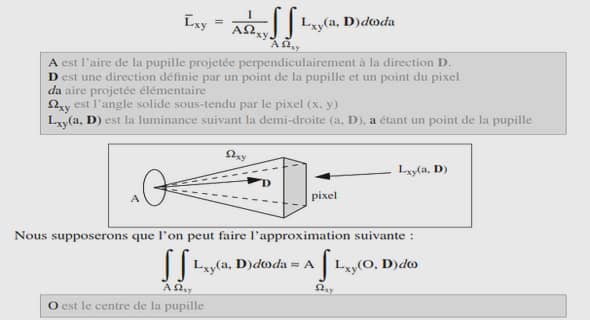
L’analyse du comportement de réseaux de neurones est un thème de recherche attractif qui attire l’attention des chercheurs venant de différentes communautés scientifiques comme les neurosciences et les neurosciences computationnelles , les systèmes dynamiques, l’informatique et plus récemment la théorie du contrôle . Ce sujet de thèse à caractère pluridisciplinaire s’inscrit dans les thématiques des neurosciences et de la théorie du contrôle. Nous nous proposons d’étudier le comportement de réseaux neuronaux, notamment les phénomènes de synchronisation du point de vue de la théorie du contrôle et de la stabilité des systèmes non linéaires.
L’analyse du phénomène de synchronisation de l’activité neuronale est motivée par l’importance de ses effets responsables de trouble de l’humeur, perte de mémoire, ou encore de dysfonctionnements du mouvement tel que le tremblement. En effet, l’activité électrique d’un groupe de neurones est à l’origine entres autres du mouvement. Le tremblement, qui est un mouvement involontaire, est le résultat d’une synchronisation neuronale pathologique. Pour pallier ce dysfonctionnement de l’activité il existe deux solutions qui sont les suivantes : le traitement médicamenteux et la stimulation cérébrale profonde. La stimulation cérébrale profonde permet de détruire l’activité synchronisée pathologique d’une zone du cerveau. Cette méthode invasive consiste à stimuler la zone du cerveau concernée par des impulsions électriques via une électrode. Le signal appliqué est réglé empiriquement sur le patient. A ce jour, il n’existe pas d’explications communément acceptées quant au fonctionnement de la stimulation cérébrale profonde permettant de rétablir une activité neuronale normale. C’est pourquoi, il est crucial d’étudier et de modéliser le réseau neuronal. L’importance et la complexité du problème font de l’analyse des systèmes neuronaux un domaine de recherche très attractif qui implique des spécialistes de communautés différentes.
Du point de vue de la théorie du contrôle, nous pouvons considérer le réseau neuronal comme un cas particulier de réseaux et de systèmes non linéaires complexes interconnectés. Pour analyser ce problème nous utiliserons donc des outils théoriques issus de la théorie du contrôle et de l’analyse qualitative des équations différentielles ordinaires. Nous nous sommes inspirés des nombreux travaux portant sur l’étude des réseaux de neurones et par les résultats sur l’analyse de la stabilité de réseaux non linéaires . Nous nous intéresserons dans ce travail à la modélisation et à l’analyse du comportement de réseaux neuronaux, notamment à l’effet de la synchronisation d’un réseau de neurones interconnectés. Le phénomène de synchronisation est très étudié dans les différents domaines des sciences et de l’ingénierie, comme par exemple en physique, en économie, en robotique pour l’étude des systèmes de puissance ou encore en neurosciences pour l’étude de l’activité neuronale. Concernant la modélisation de l’activité neuronale, nous pouvons aborder le problème de deux manières différentes. L’une d’entres elles consiste à utiliser des modèles mésoscopiques qui décrivent les dynamiques de l’activité de populations de neurones. La seconde manière de se placer est d’utiliser des modèles microscopiques qui décrivent les dynamiques de chaque neurone interconnecté et c’est ce type de modèle que nous utiliserons. Il existe deux approches pour la modélisation microscopique de l’activité neuronale. L’une de ces approches ayant un aspect plus physiologique utilise des modèles dédiés à la description des dynamiques neuronales, tels que les modèles de Hodging-Huxley [60], FitzHugh-Nagumo [49] ou encore le modèle de réseau de Hindmarsh-Rose [56, 57]. L’autre approche utilise des modèles plus abstraits qui ne sont pas basés sur la modélisation de l’aspect physiologique. Ces modèles permettent d’analyser certaines caractéristiques de l’activité, comme le modèle de Kuramoto [77], ou encore le modèle intègreet-tire .
Un neurone est une cellule nerveuse présente dans le système nerveux cérébral qui sert à traiter l’information et à communiquer. Il est composé d’un noyau, qui est à la source de son existence et qui est situé dans le corps cellulaire. Il est également constitué d’un axone, de dendrites et de terminaisons nerveuses, qui permettent de relier les neurones entres eux. Un neurone a deux propriétés fondamentales : excitabilité et conductivité. En effet, toute cellule nerveuse est caractérisée par sa capacité à réagir (cette réaction est appelée potentiel d’action) à une information reçue (le stimulus), c’est la réactivité. Le terme conductivité signifie que chaque cellule peut transmettre à son tour une information via différents modes de communication : soit par émission d’un potentiel d’action, soit par transfert chimique ou électrique . C’est par les dendrites que le neurone reçoit les informations. Les terminaisons axonales quant à elles permettent d’envoyer l’information. Ces terminaisons peuvent transmettre le message soit à une cellule nerveuse (neurone) soit à une cellule musculaire. Tous les composants d’un neurone sont délimités par la membrane cellulaire au travers de laquelle des échanges chimiques ou électriques peuvent avoir lieu . L’espace entre les membranes cellulaires est appelé espace synaptique et l’ensemble membranes plus espace synaptique forme la synapse. La membrane cellulaire permet l’échange d’ions entre le milieu intracellulaire et extracellulaire. On trouve dans ces deux milieux entre autres ions, des ions K+ (potassium) et des ions Na+ (sodium). Il n’y a pas la même concentration d’ions dans chacun des milieux. De plus, puisque ces ions sont chargés électriquement ils créent une différence de potentiel entre le milieu intracellulaire et le milieu extracellulaire (potentiel membranaire) . Le potentiel membranaire est soit à l’état de repos c’est-à dire que le potentiel est constant et est d’environ -70mV, soit dans un état transitoire durant lequel est généré un potentiel d’action (PA). Cet état transitoire est très court, de l’ordre d’1ms et est provoqué par la réception d’un message nerveux. Dans la suite nous ferons une brève description de ces deux états.
Chaque cellule nerveuse contient des ions chargés électriquement dont la concentration est différente à l’extérieur et à l’intérieur de la cellule. C’est ce qui est à l’origine de la différence de potentiel membranaire. L’échange d’ions entre le milieu extracellulaire et le milieu intracellulaire se fait par des canaux ioniques. Certains canaux sont plus perméables à un type d’ion qu’à un autre, et permettent l’échange d’une certaine concentration de ces ions dans un sens défini par le gradient électrochimique. Lors d’un stimulus la perméabilité des canaux sera modifiée laissant ainsi passer des ions K+ et des ions Na+ de part et d’autre de la membrane cellulaire.
A l’état de repos, il n’y a pas d’influx nerveux, le potentiel membranaire est d’environ -70mV . Comme nous venons de le voir, le potentiel membranaire est dû à la différence de concentration des ions (chargés électriquement) à l’intérieur et à l’extérieur de la cellule. Les canaux ioniques, à travers lesquels se font les échanges, sont d’avantage perméables aux ions K+ qu’aux ions Na+, ils laisseront donc passer plus d’ions K+ d’un milieu à l’autre. Le gradient de concentration et la concentration élevée des ions K+ à l’intérieur de la cellule entrainent une fuite de ces ions vers l’extérieur de la cellule, y apportant une charge positive . Les échanges d’ions Na+ étant moins importants, ils ne peuvent pas équilibrer le potentiel membranaire qui est alors négatif. Cet état de repos sera perturbé par un stimulus qui générera un PA.
1 Introduction |