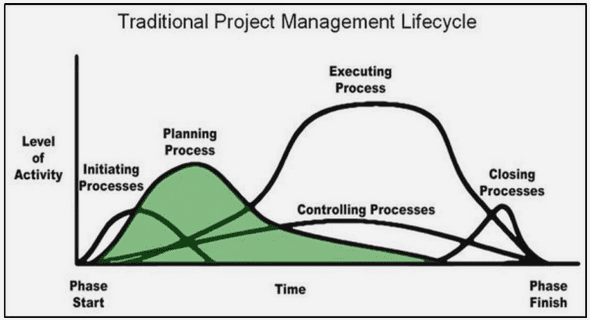
Benefits and costs of adaptation to constant and alternating polluted environments
Environmental changes are assumed to have increased in frequency and intensity throughout the world, as a result of anthropogenic activities (Millennium Ecosystem Assessment, 2005). These sudden and important changes may be extremely critical for the future of natural populations (Tilman & Lehman, 2001 ; Bell & Collins, 2008). Genetic variation is supposed to allow populations to adapt quickly to severe and novel stressors, and thus to limit their risks of extinction (Hoffmann & Parsons, 1991 ; Charlesworth & Hughes, 2000 ; Reed et al., 2003 ; Bell & Collins, 2008). Experimental and field studies have shown microevolution for populations in response of different pollutants such as xenobiotics or heavy metals (Xie & Klerks, 2003 ; Ward & Robinson, 2005 ; Morgan et al., 2007 ; Lopes et al., 2008 ; Brausch & Smith, 2009 ; Salice et al., 2010 ; Jansen et al., 2011b). Here we consider as pollutants substances that exceed a threshold concentration causing adverse effects on all or part of the ecosystem (AFNOR, 1994). Rapid adaptation goes through the selection of individuals with characteristics providing a better fitness in the given environment, and a disappearance of less fitted genotypes in the short time frame of a few generations (Hoffmann & Parsons, 1991 ; Posthuma & Van Straalen, 1993 ; Morgan et al., 2007). This adaptive response is assumed to come with a cost: the genetic impoverishment leads to a reduction of the evolutionary potential of the population, which may restrict it from dealing with novel selection pressures associated with subsequent stressors (Bergelson & Purrington, 1996 ; Coustau et al., 2000 ; Xie & Klerks, 2003 ; Salice et al., 2010). Understanding under which conditions adaptation to an environment is costly is a critical issue in evolutionary and in conservation biology. Firstly, rapid adaptation can be associated with a reduction of genetic diversity (Ward & Robinson, 2005 ; Athrey et al., 2007) generally correlated to a decrease in individuals’ fitness (Reed & Frankham, 2003). Secondly, associated to the reduction of genetic diversity, costs of adaptation can be due to selection of individuals with specific strategies conferring a selective advantage in the novel environment. The selection can be done indirectly on the life history traits. For example, the selection of genotypes allocating more resources to detoxification mechanisms at the expense of other fitness related functions (e.g. growth or reproduction). In the absence of the pollutant these highly detoxifying genotypes may show a reduced fitness compared to the lowly detoxifying genotypes (Kraaijeveld & Godfrey, 1997 ; Burdon & Thrall, 2003). There are also cases where the selection can act directly on traits, such as growth and reproduction. For example, a rapid growth and early reproduction can reduce 114 Article IV – Benefits and costs of adaptation the internal concentration of pollutant by dilution and allow some reproduction before the damages of the pollutants are too high (Sibly & Calow, 1989). However, in the absence of the pollutant these strategies may not be the most optimal. Evidences are increasing of costs of adaptation to pollutants in multigenerational experiments (Shirley & Sibly, 1999 ; Ward & Robinson, 2005 ; Wang et al., 2010 ; Jansen et al., 2011b). The occurrence of adaptation costs and their detection will depend on (i) the similarity of selection pressures between the previous and the novel environment (Travisano & Lenski, 1996 ; Jasmin & Kassen, 2007), (ii) the intensity of selection (Anderson et al., 2003). We also assume that (iii) trade-offs between the measured traits may constrain their independent evolution and thus lead to potential adaptation costs. For example, the evolution of one trait could entail a cost of adaptation on other, genetically correlated, traits (Falconer & Mackay, 1996 ; Roff, 2002b ; Roff & Fairbairn, 2007). Furthermore, fundamental trade-offs have more chance to be perceptible than trade-offs between traits less related to fitness. This is the case in the trade-off between growth rate and yield of Escherichia coli populations (Novak et al., 2006). For all these reasons, costs are not systematically found (see Coustau et al., 2000 ; Reznick et al., 2000 ; McCart et al., 2005 ; Lopes et al., 2008). Consequently, it becomes necessary to compare the effects of different (i) previous polluted environment of evolution and (ii) novel stressful environments on the existence of adaptation costs to define a comprehensive and predictive framework. Experimental studies on the costs of adaptation generally focus on costs induced by the evolution in response to a single stressor (e.g. Ward & Robinson, 2005 ; Lopes et al., 2008) or to a combination of stressors (e.g. Jasmin & Kassen, 2007 ; Jansen et al., 2011a). Comparatively, few studies on the evolution in a heterogeneous environment have been proposed yet (see Turner & Elena, 2000 ; Reed et al., 2003 for temporal heterogeneity), despite the fact that they probably reflect more natural conditions (Levins, 1968 ; Hedrick, 1974, 1976, 1986). Furthermore, environmental heterogeneity is assumed to help maintaining a higher level of genetic variation (Hedrick, 1986 ; Roff, 2002b). In such a heterogeneous environment, populations may not reach the same level of adaptation (but see Turner & Elena, 2000), but they may also suffer lower costs of adaptation than if they were evolving in a homogeneous environment (Reed et al., 2003). It is thus necessary to conduct more studies comparing the evolutionary responses of populations to constant and fluctuating selection pressures.
Population maintenance and prior environment of evolution
Because of its short life cycle, small body length, and ease of handling, C. elegans represents a good metazoan model to perform microevolution experiments (Braendle et al., 2008). To obtain a study population with a large genetic diversity, we used in this study a stock population of C. elegans composed of a mixture of 16 wild isolates (Teotónio et al., 2012). The population was kept in the experimental conditions described in Teotónio et al. (2012) for over 140 generations prior to our study. During the experiment we changed the conditions (see article III). Briefly, we placed 500 individuals in a 9 cm diameter Petri plate filled with an agar medium seeded with 1 ml UV-killed Escherichia coli OP50 strain as food source. After three days of population development we transferred 500 individuals from all developmental stages into a new Petri plate. We produced six replicated experimental populations. Generation time in C. elegans (i.e. time to complete a life cycle) is lower than three days (Byerly et al., 1976). The population was composed of males for an androdioecious breeding system (i.e. self-fertilization of hermaphrodites and facultative outcross with males). Nematodes were cultured throughout the experiment at 20°C and 80% of relative humidity.
Selection experiment
For this experiment we chose to use uranium and salt environment as stressors. Uranium is a natural radioactive heavy metal which concentrations in sediments or surface soils have increased recently as a result of human activities such as mining (UNSCEAR, 2000 ; Lottermoser et al., 2005). Exposure to natural uranium may induce both chemical and radiological effects, although uranium is assumed to have relatively higher chemotoxic than radiotoxic effects (Thomas & Liber, 2001 ; Kuhne et al., 2002 ; Miller et al., 2002 ; Zeman et al., 2008 ; Mathews et al., 2009). Salt concentration has increased recently in ecosystems, for many reasons, the most important one being the intensive irrigation of some cultivated lands (Rengasamy, 2006 ; Verwey & Vermeulen, 2011). High salt exposure is an extreme hypertonic stress that provokes a rapid water and solute content loss in cells of C. elegans (Lamitina et al., 2004). After repeating this protocol for about 40 generations, the individuals from the six replicates were mixed before being transferred into four different environmental conditions: a control environment (as described above) and three polluted environments, identical in all aspects 118 Article IV – Benefits and costs of adaptation to the control, except for the addition in the agar medium of (i) 1.1 mM of uranium (uranyl nitrate: UO2 (NO3)2, 6H2O; Sigma-Aldrich, France), (ii) 308 mM NaCl or (iii) alternating uranium and salt at each generation (salt for odd generations). For each type of environment we created six replicate populations of 500 individuals each. Thereafter we will refer to different population evolving in these environments as control, uranium, salt and alternating populations. Uranium and salt concentrations entailed a reduction of fertility about 60% at the first generation of exposition, which corresponds to potentially strong selection pressures. We previously described how we added pollutant in the agar medium (article III). Generation time, however, varied between the treatments. In particular, it was delayed in the NaCl-treatment compared to the other treatments, and therefore depending on the treatment, each experimental iteration (i.e. three days) may correspond to either a generation or a bit less than a generation. However, to simplify we kept using the term generation throughout the text. Here, we present measured of phenotypic traits at three different generations to observe the effects of treatments at the beginning (generation 1) and at the end (generation 22) of the experiment. Moreover, we decided to focus on genetic differentiations among populations and not to changes caused by withinindividual (Scheiner, 1993) or to cross-generation phenotypic (i.e. maternal effects; Räsänen & Kruuk, 2007) plastic response to the novel environment. We thus show the results starting at the fourth generation.
Common-garden and reciprocal-transplant experiments
Our objective was to estimate the costs of adaptation to uranium, salt, and the alternating treatment. Thus starting from generation 6 of the multigenerational experiment, and then every three generations, we isolated 500 individuals from each replicate to run a common-garden experiment by putting the individuals from each replicate and for each treatment back into the control environment. Differences observed in the control environment between the populations that have evolved in different prior environments can be attributed to genetic changes caused by the evolution during the multigenerational experiment (Levins, 1968 ; Conover & Schultz, 1995). At generation 18, we ran a reciprocal-transplant experiment in which samples of each replicate population from each treatment were transferred into uranium and salt environments. A negative interaction between the prior environment and the transplant environment on the trait Article IV – Benefits and costs of adaptation 119 measured should reveal some costs of adaptation to the prior environment (see below statistical analyses). Finally, at the end of the multigenerational experiment (i.e. generation 23), we realized one last common-garden experiment similar to the previous ones in all the conditions, with the exception that we placed the populations in incubators at two different temperatures: 20°C (i.e. similar to the control one in previous experiments) and 25°C. Such an increase provokes a reduction of development time and fertility and an increase of speed growth in C. elegans (Byerly et al., 1976). Additional reduction of fitness at 25°C for populations that lived previously in polluted environment compared to control populations should affect the ability to deal with an environmental stress. The costs had to be measured at least after two generations to overcome effects caused by different parental environments (Mousseau et al., 2009 ; Kawecki et al., 2012). Thus as mentioned above for generation 4 of selection experiment, prior to all measures of phenotypic traits each population spent three generations in the control or novel environment (same transfer at each generation than in the selection experiment) to ensure that the responses observed were due to the genetic differentiations.