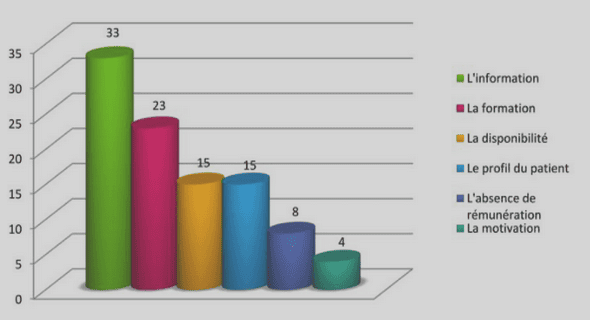
CRIBLAGE ET ÉVALUATION AGRONOMIQUE D’ACCESSIONS DE COURGETTE
Culture de la courgette
La culture de la courgette se fait au champ, sous serre ou sous tunnel. Quelque soit le type de culture utilisé, certaines pratiques culturales sont indispensables.
Choix et préparation du terrain
Pour le Choix de la parcelle, il faut envisager les sols bien drainés et respecter les distances d’isolement. Un labour pourra être effectué pour ameublir le sol, favoriser un bon enracinement et éviter les stress hydriques. Il est conseillé de travailler le sol avec des outils à dents en conditions sèches. L’objectif est d’obtenir une structure grumeleuse et aérée ainsi qu’une couche superficielle fine et humide pour une bonne germination, car la courgette a de gros cotylédons dont il faut favoriser l’émergence (Abatzian et al., 2003).
Semis
Le semis s‟effectue à partir du mois de avril. La méthode de semis dite en poquet à environ 2 ou 3 cm de profondeur avec un espace de 60 à 80 cm entre les poquets est vivement encouragée. Si le semis est réalisé en pot, il est préférable de repiquez en pleine terre 4 à 6 semaines plus tard. Cela a l’avantage de protéger les jeunes 5 plants contre les limaces jusqu’à leur bon développement (www.Jardiner malin courgette, 2012).
Humidité
On distingue 4 phases de besoin en eau au cours du cycle de la courgette. Du semis à la levée, le terrain doit être humide. Au moment du semis, un manque d’eau provoquerait une levée irrégulière. De l’installation de la culture jusqu’au stade 4 -5 feuilles un stress hydrique précoce aura des répercussions sur l’enracinement, l’homogénéité de la végétation et le nombre de fruits par plante. La courgette est très sensible au manque d’eau en début de floraison. Durant la maturation des fruits, l’irrigation n’a aucun effet (Abatzian et al., 2003).
Fertilisation
La courgette se développe bien sur les terres humides et riches en humus. L’azote joue un grand rôle dans la production de matière sèche. Quant à l’équilibre N/K, elle influe sur la formation des fleurs femelles. La plus forte demande en humus se situe au début du grossissement des fruits. Les besoins en azote de la courgette sont de l’ordre de 60 à 80 kg / ha sur tout le cycle (Abatzian et al., 2003). Pour la fumure azotée, il faut également prendre en compte le taux de minéralisation des apports organiques qui varie de 30 à 60 % selon les produits et les conditions climatiques de l’année. En ce qui concerne la fertilisation phosphopotassique on apportera entre 100 et 120 kg / ha de phosphore sous forme de phosphate naturel (sols acides), de scorie (sols acides ou neutres), de phosphate aluminocalcique (sols basiques) ou de farine d’os (tous sols) et de 100 à 120 kg/ ha de potassium (Abatzian et al., 2003).
Principales exigences de la plante
La courgette préfère les sols profonds, souples, se réchauffant rapidement mais suffisamment drainés, bien pourvus en matière organique et un pH compris entre 6 et 7,5. Un sol léger favorise la précocité. La courgette est exigeante en chaleur. Elle gèle à 0°C. La croissance de la plante est pratiquement nulle à 5°C et les températures optimales de croissance et de fructification sont de 16 à 24°C. Au-delà de 30-35°C et selon les conditions d’alimentation hydrique, la plante peut faner de manière plus ou moins irréversible. C’est une culture qui craint l’excès d’humidité et qui a besoin au moins de quelques heures par jour d’irradiation solaire (Abatzian et al., 2003). I.1.6.6. Cycle de la culture et développement Les graines germent 5–7 jours après le semis, ou plus tôt (3 jours) si l‟on fend soigneusement le tégument. Les plantes développent un vaste système racinaire fibreux et leur croissance est indéterminée. De nombreux cultivars modernes de courgette ont un port buissonnant qui se caractérise par de courts entre-nœuds et une ramification rare ou absente. La floraison, qui débute 30–40 jours après la levée de la plantule, est plus ou moins continue. La pollinisation est effectuée par les insectes, principalement les abeilles et les guêpes. Les premiers fruits immatures peuvent être récoltés 50–60 jours après la germination. Les fruits mûrs peuvent se récolter au bout de 90–100 jours. Quand les températures sont basses (10°C la nuit, 20°C le jour), des fruits parthénocarpiques peuvent se former (Abatzian et al., 2003).
Récolte et rendements
La récolte est à faire au couteau ou avec un sécateur, afin de couper le pédoncule et non pas le casser, et de préférence en conditions fraîches, dès que les fruits atteignent le calibre souhaité. La courgette est un produit normalisé pour lequel il existe 2 calibres commerciaux (14/21 et 21/26, correspondant à la longueur du fruit en cm). Le calibre 14/21 représente 80% de la production. Les courgettes de ce calibre pèsent environ 100 grammes (www. Wikipedia .Courgette, 2011). Plusieurs récoltes peuvent donc être faites par semaine en fonction du débouché. La récolte manuelle peut atteindre 50 kg/ heure / personne. Pour faciliter la récolte, il est préférable de cultiver les variétés au port aéré et au feuillage peu piquant. Il faut veiller à ne pas blesser le fruit à la récolte, son épiderme étant particulièrement fragile. La récolte peut être facilitée par l‟utilisation de remorque avec des tapis ou de petites plateformes automotrices. Le rendement moyen est de 2 à 3 kg / m² en plein champ (calibre visé = 14/21) et 4 à 5 kg/ m² sous abri (www.interbiobretagne.asso.fr, 2011). I.1.6.8. Conservation Les courgettes se conservent jusqu‟à 8 / 10 jours dans un endroit sec et ventilé, mais perdent leur brillance et leur fermeté après 4-5 jours. Les conditions idéales de conservation au frigo sont entre 0 à 4 °C avec une hygrométrie de 85-90% (w www.interbiobretagne.asso.fr, 2011).
Production de semences
La production de semences est possible à condition de respecter quelques règles, notamment en matière de distance d’isolement avec d’autres parcelles (maraîchères mais aussi potagères privées) où sont cultivées des courges (Pâtisson, citrouille…). La distance minimale d’isolement est de 1000 m entre 2 populations de courgettes, 2000 m entre courgettes et autres espèces de courges du genre (ou variété ?) Pepo. Bien que la courgette soit une espèce fondamentalement allogame, il est également possible de multiplier des lignées pures (par autofécondation) de courgette car il y a peu de dépression génétique chez cette espèce. La semence est mûre 70 jours après fécondation. Un fruit donne de 130 à 180 graines, le PMG (Poids de Mille Grains) de la courgette est compris entre 130 et 150 g. Les semences ont une capacité germinative d’environ 3 ans (MAEP). Récoltés, les fruits peuvent se conserver 2 mois avant d’en extraire les graines. Par contre, les courgettes doivent être extrêmement saines, car quelques fruits pourris peuvent rapidement compromettre une partie de la récolte. Le séchage est une étape très importante pour les semences. Il peut être réalisé à l’air ambiant lors des premières récoltes. Quand les semences ne sont pas bien séchées, des champignons saprophytes peuvent alors se développer et altérer au moins la présentation du lot (Abatzian et al., 2003).
Rotation culturale
Etant très exigeante en matière organique, la courgette sera placée en tête de rotation ou après un engrais vert. Bien qu’exigeante, la courgette n’épuise pas trop les sols. Une succession trop importante de courgettes sur la même parcelle (moins de 3 ans entre 2 cultures) pourrait entraîner des problèmes sanitaires (ex : nématodes sous abri). La culture de courgette succède les légumes feuilles (chou, Laitue, Épinard, Oseille) et précède les légumes racines (carotte, navet, manioc, 7 radis, betterave). Il est déconseillé de pratiquer une succession des légumes fruits tels que le concombre, la courgette, le haricot, le pois, la tomate, etc. (F.A.O, 1997).
Utilisation
La courgette se mange cuite ou crue comme le concombre. Elle est consommée sous forme de bouillie et accompagnée d’autres légumes, comme dans la ratatouille, sautée, frite, farcie, en gratin ou en soupe. Dans la soupe, elle remplace la pomme de terre. Les fleurs mâles stériles sont enlevées tôt pour ne pas freiner la fructification et sont consommées, farcies ou en beignet, notamment dans les Balkans, en Italie, dans la région niçoise. La fleur de courgette farcie est une spécialité de la cuisine du Vietnam. Les graines ou la pulpe de Cucurbita pepo sont comestibles, de même que celles d’autres cucurbitacées, crues ou grillées. Par exemple la poudre de graines de citrouille s’emploie en Chine et aux Etats-Unis comme ingrédient des sauces à salade, ainsi qu’en pâtisserie. L’huile des graines s’utilise comme huile d’assaisonnement en Europe, et en Inde elle sert pour la cuisson. En Afrique, la pulpe est utilisée comme cataplasme pour traiter les brûlures et les inflammations, ainsi qu’en compresse rafraîchissante pour traiter maux de tête et névralgies. Elle est aussi appliquée sur des tumeurs et des cors (Messiaen & Fagbayide, 2004). Au Sénégal, elle est surtout utilisée par la population d’origine libanaise et les européens à la place du concombre ou dans la soupe.
Ennemis de la courgette
Parmi les causes de réduction significative des rendements de la courgette figurent les ravageurs et les maladies biotiques (dont les viroses). I.1.7.1. Ravageurs Les principaux ravageurs animaux de la culture de courgette sont d’abord les acariens (avec l’espèce Tetranychus urticae) qui posent peu de problèmes, sauf parfois sous abri. Ils se développent par temps chaud et sec. Un bassinage (aspersion) de la culture permet de réduire leur impact. Enfin les Pucerons (Aphis gossypii, Myzus percicae), vecteurs de virus, qui affaiblissent les cultures. En culture sous abri, ou par forte chaleur, les foyers peuvent se développer très rapidement et favoriser le développement de fumagine (champignon noir, qui colonise le feuillage et nuit à la photosynthèse) (www.interbiobretagne.asso.fr, 2011).
Maladies biotiques
Parmi les maladies causées par les champignons ont peut citer trois exemples. D‟abord l‟oïdium (Erysiphe cichoracearum – Sphaerotheca fulginea), favorisé par des nuits fraîches, des températures élevées dans la journée (26 °C) et une forte hygrométrie. Contrairement aux autres champignons, l’oïdium n’a pas besoin de film d’eau pour se développer. Ensuite nous avons la pourriture molle des fruits causée par Botrytis cinerea, pourriture grise à l’extrémité des fruits avant la chute de la fleur ou sur les fruits avortés. Enfin la pourriture blanche due à Sclerotinia sclerotiorum, qui provoque un épais feutrage blanc sur les fruits. Ces 2 dernières maladies sont favorisées par une forte hygrométrie (www.interbiobretagne.asso.fr, 2011). Les maladies virales ont elles aussi une grande incidence économique. Le virus de la mosaïque du concombre (CMV) et le virus de la mosaïque jaune de la courgette (ZYMV) provoquent des mosaïques déformantes sur les fruits et sur les feuilles en 8 commençant par l’apex et sont capable de dévaster des cultures. Ces virus sont transmis par les pucerons (www.interbiobretagne.asso.fr, 2011).
Virus de la mosaïque jaune de la courgette(ZYMV)
Historique et répartition géographique
Le Zucchini yellow mosaic potyvirus (ZYMV) est isolé pour la première fois en Italie en 1973 et décrit en 1981. Par la suite ce virus est identifié dans tous les continents au cours d’une décennie. C‟est l’un des virus les plus économiquement importants des cucurbitacées (Desbiez & Lecoq, 2003). Le virus de la mosaïque jaune de la courgette (ZYMV) se trouve dans de nombreuses régions du monde où les cucurbitacées sont cultivées. En Afrique, le ZYMV a été enregistré en Algérie, en Egypte, à l‟Ile Maurice, au Liban, au Maroc, au Kenya et en Zambie. Il a été aussi rapporté en France, en Allemagne, en Italie, en Israël, en Espagne, aux Etats-Unis, en Australie, au Royaume-Uni et en Floride (Lisa & Lecoq, 1984, Nono-Womdim, 2001).
Morphologie et cycle de vie
Le virus de la mosaïque jaune de la courgette est un virus avec des particules filamenteuses flexueuses d’environ 750 nm composé de l’ARN simple brin et une protéine d’enveloppe (Lisa & Lecoq, 1984). Pour que le cycle viral puisse débuter, il faut qu‟une particule virale pénètre à l‟intérieur d‟une cellule végétale. Puis le virus utilise la machinerie cellulaire de la plante pour proliférer. La présence de micro-blessures souvent créées par des vecteurs (insectes piqueurs) est requise pour permettre la pénétration des virus. Il débute par la libération de l‟acide nucléique dans le cytoplasme: c‟est la décapsidation. Le pH intracellulaire légèrement basique déstabilise suffisamment le virus pour que l‟acide nucléique porteur de l‟information génétique soit libéré et devienne accessible. La décapsidation et la traduction sont souvent liées. En effet, les ribosomes de la cellule se fixent à l’extrémité 5′ de l’ARN viral accessible à l’extrémité de la nucléocapside. En se déplaçant le long de l’ARN pour la traduction, ils dissocient les sous unités protéiques de la capside. Les premières protéines synthétisées sont tout de suite utilisées et interviennent dans la réplication (Costa, 2003). La réplication de l’ARN (+) viral s’effectue en deux étapes. Dans un premier temps, la réplicase se fixe en 3′ de l’ARN génomique puis synthétise un ARN de polarité opposée (-), complémentaire de l’ARN viral. Ce dernier va servir de matrice à la réplicase virale pour la synthèse d‟un grand nombre de molécules d‟ARN (+). La traduction de ces copies conduit à l‟accumulation de protéines virales en particulier la protéine de capside, permettant la formation de nouveaux virions par autoassemblage (jusqu‟à 106 particules virales par cellule). Mais l’encapsidation du génome viral est encore un phénomène mal connu. Une fois le virus multiplié dans la cellule, il va infecter les cellules adjacentes en empruntant des canaux préexistants qui relient les cellules entre elles, les plasmodesmes. Ce mouvement du virus de cellule à cellule dit de courte distance, s’explique par le fait que les virus ne peuvent se libérer des cellules par bourgeonnement en raison de la présence de la paroi pecto-cellulosique. Les plasmodesmes ont une structure interne tellement complexe qu’ils ne laissent passer que des molécules plus petites que les virus. Pour passer d’une cellule à l’autre, le virus nécessite l’intervention d’une ou plusieurs protéines virales dites protéines de mouvement. Grâce au mouvement à courte distance, le virus gagne petit à petit le système vasculaire et plus précisément les cellules du 9 phloème, impliqué dans la circulation de la sève élaborée. Le virus peut alors se propager dans la plante entière et produire une infection généralisée dite aussi infection systémique. On parle alors de mouvement à longue distance (Costa, 2003)
Introduction |