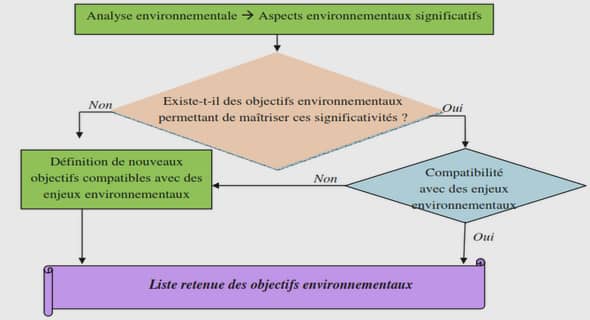
SAMPLING DESIGN
For this study, we used a part of the forest inventory database of the ministère des Ressources naturelles du Québec that covers two administrative regions (Abitibi-Témiscamingue and Nord du Québec). The sampling units of the forest inventory database was a circular plots of 0.04 ha, distributed every 150 rn along transects of at !east 1.5 km. Transects were first randomly determined and selected for composition by the Direction des inventaires forestiers of the ministère des Ressources naturelles du Québec (Forêt Québec, 2000). Starting with more than 8000 plots located over 40 000 km2 , we selected all plots (N=43) that were on lacustrine clay deposit, bad moderate to imperfect drainage and contained aspen and black spruce (Figure 1.1 ). Plots having more than 10% of the ir total basal area occupied by species other than aspen or black spruce were also rejected of the forest inventory database. To ensure that drainage conditions were not associated with the abundance of aspen and that the two factors were not mixed up, we performed at test to compare the relative basal area of aspen between plots with moderate drainage and ones with imperfect drainage.
This test revealed no statistically significant difference (t value: 0.14; p = 0.8905). The average aspen relative basal area was 27% for imperfectly drained sites and 26% for moderately welldrained sites (aspen relative basal area was determined by the ratio of aspen basal area on total basal area of the stand). For the 43 plots selected, the mean density was 44 75 stems/ha varying from 1200 stems/ha to 12 425 stems/ha and, the mean age of dominant black spruce trees was 55 years varying from 23 to 75 years (Table 1.1). In general, selected plots are relatively young mixed stands of black spruce and as pen on well-drained soi!. Diameter at breast height ( dbh) was measured for every stem in each plot. In each plot, the height of three dominant stems of black spruce was measured with a clinometer and the age was assessed by coring. Tree merchantable volume was calculated with equations using dbh and an estimated height to calculate the volume of each stem of> 10 cm dbh (Perron, 1985). These equations are inadequate for small stems under 10 cm of dbh and the information required to calculate the volume of these stems was not available in the forest inventory database thus total gross volume could not be calculated.
Individual species scale
In order to explore the relationships between aspen relative basal area and black spruce average dbh without the influence of stem density, a polynomial regression analysis was performed between the relative basal area of aspen and the residuals of a regression analysis between black spruce average dbh and stem density. Polynomial regression was performed as a multiple regression with the relative basal area of aspen and the square of relative basal area of aspen as independent variables. Despite the common assumption that dominant tree height is not influenced by stand density, we used a polynomial regression analysis for investigating effects of aspen on black spruce average dominant height with density as a covariable. With a proportion of aspen basal area greater than approximately 45%, a negative influence of aspen was observed on the residuals of both regression analyses between dbh and height of black spruce and stem density. A piecewise regression was performed to test the significance of the model with two segments and to identify a breakpoint.
The following equations were fit to the data: ifx<‘t theny=bo+b1x if x::=:: ‘t then y= bo +(bi+ bz)x- hz• where x is relative basal area of aspen, ‘t is the breakpoint, b0 is the y-intercept, b1 is the slope for the first segment and ~+b2 is the slope for the second segment (Seber and Wild, 1989). The two piece regression was non-linear and involved parameter estimation (nlin procedure, Version 8, SAS Institute lnc. 1999-2001). Since the final parameter values were dependent on initial parameters, data were first assessed visually to estimate parameters. Regressions were considered significant when the 95% confidence intervals of one or more consecutive slopes did not contain zero (p < 0.05). According to the breakpoint identified by the piecewise regression, it was then decided to test the influence of aspen on the residuals of black spruce average dbh and dominant height on a restricted gradient of aspen basal area (from 1% to 41% of the total stand basal area).
A linear regression analysis was performed between the relative basal area of aspen and the residuals of a regression analysis between black spruce average dbh and stem density on a restricted gradient of aspen basal area (from 1% to 41% of the total stand basal area). The same method was followed to test the relationship between the relative basal area of aspen and mean dominant height of black spruce to control for the influence of stand density on a restricted gradient ofaspen basal area (from 1% to 41% ofthe total stand basal area). Finally, an analysis of covariance, with stem density as a covariable, was used to test the influence of aspen on black spruce stem volume. This analysis was performed on two types of sites with different proportions of aspen: 0-5% of total basal area, which were considered as control sites, and 5-15% of total basal area, which were considered as sites with a slight presence of aspen. Because the assumptions of the covariance analysis were not met, a nonparametric covariance analysis was performed on four ranges of aspen relative basal area (0-5%, 5-15%, 15-45%, 45-100% of basal area), and another nonparametric covariance analysis was performed on three ranges (0-5%, 5-15%, 15-45%) but there was no significant difference between these categories.
DISCUSSION
Despite the presence of similar abiotic conditions, black spruce growth differed with the proportion of aspen in the stand and the nature of the influence of aspen also changed with the proportion of aspen in the stand. The apparent absence in the literature (see review by Rothe and Binkley, 2001) of a clear trend regarding the effect of mixed stand management on stand productivity could be explained in part by the variability in the influence of the companion species with respect to its abundance in a stand. In this study, the presence of as pen in proportions lower than 41% of total basal area of the stand increased black spruce dbh and height. According to the piecewise analysis models, we estimated an increase in dbh of approximately 13 to 17 cm, and an increase in height of 12 to 17 rn with aspen varying from more than 0 to 41% of relative basal area, which suggests that facilitative production mechanisms could be present between aspen and black spruce. Reported site indices for black spruce in Quebec’s commercial forest vary from 9 to 18 meters (Pothier and Savard, 1998). Observed gains are therefore highly significant from a growth and yield perspective. For example, the commercial volume of a 90-year-old medium density black spruce with a site index of 12 rn is 80m3/ha compared to 240m3/ha for a similar stand with a site index of 18 rn (Pothier and Savard, 1998).
In addition to a positive influence of aspen on diameter growth of black ~ruee, which could increase the economie value per hectare, the presence of 5 to 15% of aspen basal area in a stand is sufficient to obtain a higher volume of black spruce per stem, which could also benefit commercial users of the forest. However, the presence of aspen seemed to negatively influence black spruce dbh (12 cm at 85% of aspen basal area) and height (13 rn at 85% of aspen basal area) when the proportion of aspen reached values greater than 41% of the total stand basal area. Over this threshold, the decrease in black spruce growth could be explained by higher interspecific competition. However, the estimation of the difference of dbh ( 1 cm) and height (1 rn) between both extremes of the gradient of aspen (more than 0 vs 85%) could be negligible. The relative proportion of aspen in the stand is a major parameter in mixed management and more studies are needed to understand the dynamics of these stands and to fix a range or a threshold of aspen proportion for successful management.
Despite the attention given to selecting plots with similar abiotic conditions, there is still a possibility that the positive influence of aspen on height and dbh was confused with differences in soil properties that were correlated with stand composition and productivity. The influence of stem density on black spruce dominant height was not expected, but this relationship has been observed before in black spruce stands of high density (Grondin et al., 2000). At the stand scale, the presence of aspen in proportions lower than 41% of total stand basal area increased the total merchantable volume of the stand. The absence of a significant relationship between the merchantable volume of black spruce and the relative basal area of aspen implies that volume gain in the stand is essentially aspen fibre. This suggests that aspen uses a different ecological niche than black spruce, thus reducing the competition between the species. In additional to providing an « extra cubic meter » of aspen fibre, the presence of aspen appears to provide similar merchantable volume of black spruce that is distributed in fewer stems of greater height and dbh.
Sampling design
Three sites were chosen according to the following criterias: gentle slope, moderate drainage, soil type and stand composition dominated by Picea mariana with heterogeneous presence of Populus tremuloides. Ali stands originate from fires that took place around 1920 according to the fire map producted by Bergeron et al. (2004). More precisely, sites 1, 2 and 3 originate from 1926, 1916 and 1916 fires, respectively, according to local data obtained from crosssectional tree-discs of 10 Populus tremuloides trees taken at ground leve! (0 rn) at each site. The sampling unit of this study was a circular plot of 14 rn in diameter, distributed at every 20 rn along a transect of at !east 180 m. The number and length of transect lin es depended on the size and form of the Picea mariana-Populus tremuloides stands. Each sampling unit was considered independent because tree species composition influences sail properties at a small spatial scale (Boettcher and Kalisz, 1991; Rhoades, 1997; Turner and Franz, 1985). Sites 1, 2 and 3 had respectively 28, 33 and 34 plots. Sites were different on the basis of stand density and sail texture (Table 2.1 ). Site 1 had higher mean stand stem density, with 3220 stems/ha, and sites 2 and 3 had mean stem densities of 2014 and 1993 stems/ha, respectively. On each site, black spruce basal area decreases as aspen basal area increases. Fine roots were mostly located at the interface between the A mineral horizon and the humus layer. Humus layer thickness was lower in site 1 and higher in site 3 (Table 2.2). To ensure that mineral sail properties were not correlated with Populus tremuloides presence, we selected five plots in each site corresponding to of the three following categories for further analyses (total of 45 selected plots): 1) plots dominated by Picea mariana with Jess than 15% of relative basal area of Populus tremuloides, 2) plots with 15 to 50% relative Populus tremuloides basal area, and 3) plots with 50 to 75 % relative Populus tremuloides basal area.
REMERCIEMENTS |