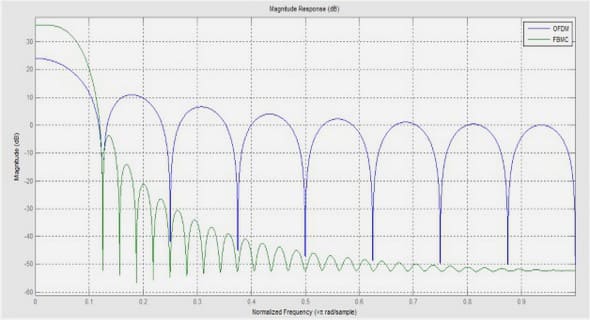
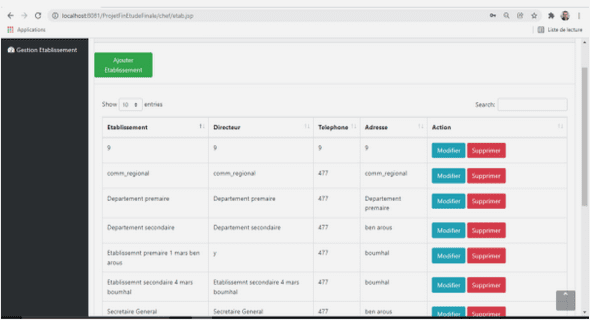
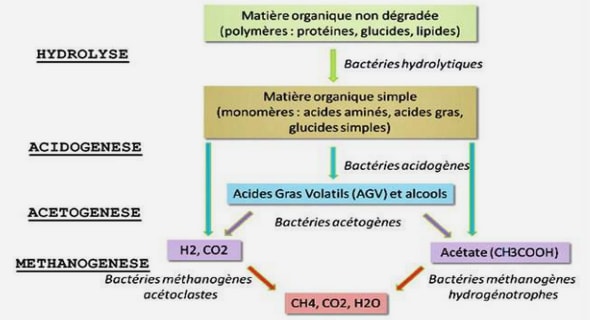
Fragmentation de l’habitat
Fragmentation et perte d’habitat
La fragmentation de l’habitat est l’un des sujets en écologie les plus étudiés depuis les 20 dernières années , du fait de la progression constante des perturbations humaines imposées aux paysages. Le terme fragmentation a souvent été confondant dans la littérature scientifique, référant à la fois à la perte d’habitat (réduction de la quantité d’habitat) et à la configuration spatiale des habitats résiduels (superficie et isolement des parcelles d’habitat). Pourtant, de nombreux auteurs considèrent la perte d’habitat et la fragmentation comme deux phénomènes distincts (Fahrig 1997, Haila 1999, McGarigal et Cushrnan 2002, Lindemnayer et Fischer 2007). Il est maintenant largement accepté que la fragmentation réfère strictement aux changements dans la configuration spatiale d’un habitat donné (rupture de la continuité), dont les effets ne deviennent importants qu’au delà d’un certain niveau de perte d’habitat (Villard 2002).
Le concept de fragmentation de l’habitat origine de la théorie de la<< biogéographie insulaire >>(MacArthur et Wilson 1967), mais a été ensuite repris et adapté pour des paysages terrestres moins contrastés (voir Haila 2002). Malgré cette littérature grandissante, il est difficile de tirer des principes universels sur les effets de la perte et de la fragmentation de l’habitat car les milieux naturels sont fragmentés à différents niveaux, ce qui complique les comparaisons (Schrniegelow et M6nkk6nen 2002). De plus, ils sont soumis à des changements continus dus aux processus naturels, ce qui rend les conclusions des études temporellement obsolètes (Haila 2002). Par ailleurs, les différents organismes réagissent de façon différente aux divers degrés de fragmentation, au contexte environnemental particulier et à des échelles spatiales et temporelles différentes (Haila 2002).
Fragmentation et échelles spatiales
Les effets de la perte et de la fragmentation de 1 ‘habitat ont souvent été évalués à 1’échelle du paysage, car ils sont souvent étudiés dans un contexte de conservation et d’évaluation des risques d’extinction de certaines espèces ou populations. L’échelle du paysage ne présente pas de définition unique, elle est plutôt relative à l’organisme ou au processus étudiés (Turner et al. 2001). L’étendue spatiale et temporelle de l’échelle du paysage n’est donc pas fixe. De plus, contrairement à d’autres branches de l’écologie, l’écologie du paysage considère une hétérogénéité spatiale et temporelle intrinsèque à son objet d’étude (Forman et Godron 1986). La <<théorie de la percolation>> prévoit que la continuité d’un habitat ne sera rompue qu’après 40% de perte d’habitat (Gardner et al. 1987). Les parcelles d’habitat ne commenceront à être isolées qu’après 60% de perte d’habitat, mais c’est au-delà de 80% de perte d’habitat que les effets d’isolement seront réellement importants. Andrén (1994) a, quant à lui, prévu la réponse des populations animales le long de ce gradient de perte d’habitat à partir d’une méta-analyse. Selon lui, la réduction de la taille des populations sera d’abord proportionnelle à la perte d’habitat puis, sous un certain seuil (au-delà de 70% de perte d’habitat), le déclin deviendra plus drastique dû à l’effet additif de la fragmentation . Cependant, la sensibilité à la fragmentation de 1 ‘habitat est spécifique à chaque espèce, selon sa capacité de dispersion, ses spécificités en termes d’habitat et l’échelle spatiale qui la caractérise (With et Cris! 1995, Andrén et al. 1997, M6nkk6nen et Reunanen 1999). Ainsi, le seuil proposé par Andrén (1994) ne peut être utilisé comme ligne directrice en matière d’aménagement (M6nkk6nen et Reunanen 1999). Les principaux effets de la fragmentation peuvent se résumer à une réduction de la superficie des îlots de forêt résiduelle, un plus grand isolement de ceux-ci et une augmentation de la quantité de lisières (Hunter 1990, Fahrig et Merriam 1994, Forman 1995). Les plus petits îlots abritent moins d’individus, le risque d’extinction y est donc plus élevé et le taux de colonisation plus faible (Fahrig et Merriam 1994).
Les effets de la perte et de la fragmentation de 1 ‘habitat peuvent se faire sentir à différentes échelles spatiales, à l’échelle du paysage sur les populations, mais aussi à des échelles plus fines qui affectent les individus et leurs comportements. Ces diverses échelles reflètent les échelles de perception que les animaux ont de leur environnement. En effet, Johnson (1980) décrit les différents niveaux de sélection d’habitat : le 1″ niveau caractérise l’aire de répartition géographique de l’espèce, le 2′ niveau détennine le domaine vital d’un individu ou d’un groupe social, le 3′ niveau définit les types d’habitats utilisés à l’intérieur des domaines vitaux et finalement le 4’ niveau réfère au micro-site ou à des attributs spécifiques de l’habitat. Ainsi, si la qualité d’un habitat diminue ou si la superficie d’une parcelle d’habitat rétrécit, un individu peut répondre à cette perturbation en incluant des habitats adjacents, parfois sous-optimaux, ou d’autres parcelles du même habitat localisées à proximité (<<théorie de la supplémentation >>, Dunning et al. 1992). Ceci aura donc des conséquences à la fois sur la superficie du domaine vital et sur les types d’habitats utilisés à 1 ‘intérieur de celui-ci (niveau 2 et 3 de Johnson 1980). À une échelle encore plus fine, les individus peuvent ajuster leurs déplacements en réponse à la perte d’habitat en évitant des milieux inadéquats (par exemple où le risque de prédation s’avère trop important). Dans ce cas, c’est la connectivité entre les différents habitats qui déterminera l’utilisation des parcelles d’habitat. Or, la perception de la connectivité est spécifique à chaque espèce, selon leurs capacités de déplacement notamment (With et Cris! 1995). Afin d’avoir un portrait complet des effets de la perte et de la fragmentation de 1 ‘habitat sur une espèce donnée, il apparaît donc important d’étudier un gradient de perte d’habitat le plus large possible afin d’inclure les effets additifs de la fragmentation et d’analyser la réponse de la dite espèce à plusieurs échelles (populations, individus, mouvements).
Le cas de la forêt boréale
La forêt boréale a de tout temps été dynamique. En effet, les feux de forêts, plus ou moins fréquents, plus ou moins intenses, et couvrant de plus ou moins grandes superficies ont toujours induit des changements récurrents du couvert forestier dans le paysage (Gauthier et al. 1996, Bergeron et al. 1998, Bergeron et al. 2004). Ainsi, la forêt boréale subit périodiquement des pertes d’habitat à l’échelle locale et régionale de même qu’elle se fragmente naturellement. La distribution des classes d’âge des peuplements forestiers créée par les perturbations naturelles suit une courbe exponentielle négative , les territoires sous régimes naturels de feux comportant une proportion importante de vieux peuplements. Ainsi, on sait par exemple que plus de 50% des forêts dépassent 100 ans (Harper et al. 2002) dans un paysage où le cycle de feux est d’environ 140 ans (Bergeron et al. 2001). Cependant, dans les dernières décennies, les coupes forestières ont ouvert de plus en plus la matrice forestière et les niveaux de perte et de fragmentation de 1 ‘habitat atteints se situent maintenant en dehors de la plage de variabilité naturelle que la forêt boréale de l’est de l’Amérique du Nord ait jamais connue (Cyr et al. 2009). L’exploitation forestière a même dépassé les feux comme principal agent perturbateur à grande échelle, et ce, dans différents secteurs de la forêt boréale (Schroeder et Perera 2002, Perron 2003, Drapeau et al. 2009). La distribution des classes d’âge des peuplements forestiers issue d’un régime équien de coupes forestières est très différente de la forêt naturelle et tronque la portion des vieux peuplements qui sont ciblés par l’industrie. Les espèces inféodées aux vieilles forêts ou à des attributs d’habitat présents dans les vieilles forêts (par exemple, le bois mort) sont donc à risque, comme c’est le cas actuellement en Fenno-Scandinavie (Berg et al. 1994).
Malgré ce constat, le contexte particulier créé par les coupes forestières est rarement considéré dans les théories sur la perte et la fragmentation de 1 ‘habitat (Boulin et Hebert 2002), car on ne se situe pas dans un cas extrême de conversion des terres, comme c’est le cas de l’agriculture ou de l’urbanisation (Forman et Godron 1986). Les paysages résultants sont plutôt caractérisés par 1) une perte temporaire de l’habitat forestier, car les parterres de coupes régénèrent en forêt (M6nkk6nen et Reunanen 1999), 2) l’importante matrice forestière environnante composée de l’habitat original (Schrniegelow et al. 1997, Drapeau et al. 2000) peut encore jouer un rôle de <<source >>pour les populations locales (Pulliam 1988), du moins pour le moment, et 3) 1 ‘habitat original résiduel est encore fortement connecté grâce à des corridors forestiers entre les parterres de coupes (Boulin et Hebert 2002).
INTRODUCTION |