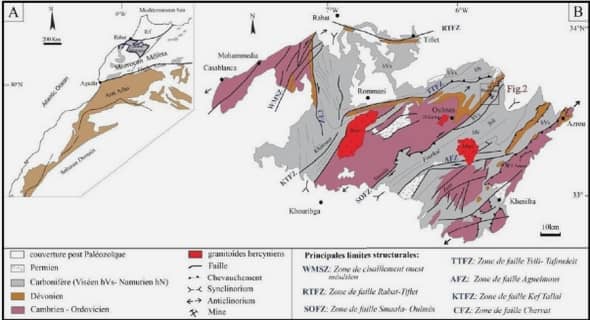
Après un long débat historique sur l’impact de l’environnement sur les individus, il est maintenant accepté que l’environnement influence au moins la répartition géographique des individus et plus généralement des espèces. En effet, les modifications d’habitat sont connues pour entraîner un changement dans leur distribution (Parmesan 2006; Parmesan & Yohe 2003; Peñuelas & Boada 2003; Walther et al. 2002). Intégrer l’évolution du paysage dans lequel vivent les individus est devenu capital pour mieux étudier et comprendre les populations naturelles. Dans cette optique, Manel et al. ont proposé en 2003 de combiner l’écologie du paysage avec la génétique des populations : les flux géniques et les adaptations locales pourraient ainsi être expliqués via des phénomènes d’ordre environnementaux. Alors qu’aucun outil ne permettait de lier ces deux disciplines aux XIXème et XXieme siècles, c’est l’évolution parallèle et progressive de la géo localisation et de la génétique qui a rendu possible leur rencontre. Une succession d’évènements va rapprocher progressivement ces disciplines pour en générer une nouvelle, appelée ‘landscape genetic’ (i.e. génétique du paysage). Etudier l’influence du paysage sur la distribution de la variabilité génétique devient alors possible.
Voici brièvement résumé l’histoire de la naissance de la génétique du paysage.
Les années 1950 ont marqué très fortement la génétique et la géo-localisation. En effet, Watson et Crick en 1953 décrivent pour la première fois la structure hélicoïdale de la molécule d’ADN. Crick obtient quelques années plus tard le prix Nobel de médecine (1962). Quatre ans après la découverte de Watson et Crick, en 1957, l’Homme entre dans l’ère spatiale avec les Russes qui envoient pour la première fois un satellite dans l’espace, son nom : Spoutnik (du russe спутник signifiant « compagnon de route »).
Plus tard, au début des années 1970, en pleine guerre froide, le département de la défense des Etats-Unis finance un projet strictement militaire visant à géolocaliser un point sur la planète en temps réel, le nom de ce projet : Navstar Global Positioning System (1973). C’est l’acte de naissance de la notion de géolocalisation. Les années 1970 marquent également la génétique avec l’apparition de deux méthodes de séquençage de l’ADN : la méthode de Sanger et celle de Maxam et Gilbert. En 1994, la constellation de satellites accueille sa 24ème unité qui rend alors possible la géo-localisation d’un point n’importe où sur la planète Terre. Les années 1990 voient également la naissance de logiciels spécialisés dans le traitement des données génétiques pour l’étude des populations naturelles. C’est ce qui deviendra la première génération de logiciels de génétique. Ces logiciels, munis d’une interface graphique, calculent un grand nombre d’indices génétiques développés au cours du dernier siècle (indice de Nei, de Wright par exemple). En 1993, Tautz définit des classes de petites séquences d’ADN répétées en tandem, c’est l’acte de naissance des microsatellites (Tautz 1993).
A partir de 1995, les progrès s’accélèrent et le lien entre géo-localisation et génétique devient de plus en plus étroit. En effet, en 2000, le président des Etats unis d’Amérique, Bill Clinton, supprime la SA (Selective Availability) qui réservait l’accès à la constellation de satellites uniquement aux forces armées. Ainsi, à partir de 2000, la constellation satellitaire est accessible à tous et des appareils de géo localisation commencent à apparaître sur les marchés (avec une précision horizontale de l’ordre de 100m). Bien que déjà abordée auparavant, c’est en 2001 que la 3ème dimension voit le jour et on passe alors à une précision de 4m horizontalement et de 8m verticalement. Les logiciels permettant de traiter ces informations de géo-localisation évoluent de façon concomitante et ESRI ArcGIS version 8.1 apparaît. Cette nouvelle version du logiciel permet d’intégrer des données en 3 dimensions, on parle alors de ‘Modèle Numérique de Terrain’. Ces avancées en informatique favorisent également l’évolution et l’expansion des logiciels de génétique. En effet, au début des années 2000, le développement de l’Internet et l’apparition d’ordinateurs de plus en plus ‘puissants’ permettent, d’une part, d’accéder plus facilement aux logiciels et d’autre part de lancer plus d’analyses en moins de temps ! Ainsi, la ‘première génération’ de logiciel de génétique (comme par exemple GENEPOP (Raymond & Rousset 1995), GENETIX (Belkhir et al. 1996), BOTTLENECK (Cornuet & Luikart 1996)) est peu à peu remplacée par la ‘deuxième génération’ essentiellement basée sur les analyses bayésiennes (par exemple: STRUCTURE (Pritchard et al. 2000) (voir figure ci –contre), BAYESASS (Wilson & Rannala 2003), DISTRUCT (Rosenberg 2004)). Ces analyses bayésiennes apparaissent comme une révolution dans les études de génétique car elles permettent d’incorporer des ‘priors’ ou a priori dans les analyses et de passer d’une échelle populationnelle à une échelle individuelle, permettant des analyses plus fines (Beaumont & Rannala 2004; Shoemaker et al. 1999).
Au début des années 2000, la géo-localisation n’est pas aussi accessible qu’aujourd’hui, tant d’un point de vu conceptuel que d’un point de vue financier. Ainsi, ce n’est que depuis 2005 que les appareils de géo-localisation (ou GPS pour Global Positioning System) se répandent dans les collectivités et les foyers. L’année 2004 voit également apparaître Google Earth et WorldWind qui permettent certes de survoler la Terre et de zoomer sur un lieu de son choix mais également d’ajouter ses propres données sur des fonds de cartes disponibles en libre accès. L’accès ‘facile’ aux appareils GPS et aux fonds de cartes numériques apporte aux études de terrain, un géo-référencement précis de chaque échantillon collecté. L’échantillon n’est ainsi plus rattaché à une population englobant un nuage de points, mais est considéré comme une entité à part entière, ayant sa propre position géographique (longitude, latitude).
La géo-localisation précise de chaque échantillon et les ressources informatiques grandissantes donne naissance à une troisième génération de logiciels de génétique prenant à la fois en compte les coordonnées géographiques et les caractéristiques génétiques de chaque individu. Parmi cette nouvelle génération de logiciels, nous pouvons citer : ALLELE IN SPACE (Miller 2005), GENALEX (Peakall & Smouse 2006), TESS (Chen et al. 2007; Guillot 2009). Ainsi, avec cette nouvelle génération de logiciels, chaque échantillon est considéré comme une entité ayant sa propre histoire et ses propres caractéristiques génétiques et géographiques.
Conséquence directe des progrès du XXème siècle, la succession de découvertes a conduit à la rencontre entre la géo-localisation et la génétique et, par extrapolation, à la jonction entre les méthodes de SIG et celles de génétique des populations. Le fruit de cette association conduità la conception de nouveaux logiciels, qualifiés dans cette thèse, de ‘troisième génération’.
Même si les logiciels de génétique et les avancées en informatique ne cessent d’évoluer chaque jour, c’est sans doute la biologie moléculaire qui révolutionnera prochainement les logiciels de génétique en faisant probablement apparaître une quatrième génération de logiciels. En effet, la plupart des logiciels de 1ère, 2ème et 3ème générations utilisent des données basées sur des ‘petites’ séquences génomiques. Aujourd’hui, nous rentrons dans l’ère de la génomique et le séquençage massif se développe de plus en plus. Pour un coût relativement modeste (2500 €) et en seulement 3 jours, il est possible aujourd’hui de séquencer 100 mégabases. C’est également la ‘course aux microsatellites’ : si 6 à 7 marqueurs microsatellites suffisaient en 2004 pour valider une étude de génétique ; il faut aujourd’hui, 12-17 marqueurs microsatellites pour être quelque peu ‘crédible’ et pouvoir prétendre à la publication des résultats. Ces marqueurs microsatellites qui dans le passé avaient remplacé les marqueurs RAPD (Random Amplified Polymorphic DNA) et qui eurent leur apogée en 2006, sont progressivement remplacés par les SNPs (Single Nucleotide Polymorphisms).
Nous sommes actuellement dans une époque charnière où la biologie moléculaire connaît une progression spectaculaire. Il ne serait donc pas étonnant de voir apparaître d’ici peu des logiciels de ‘quatrième génération’ intégrant à la fois les données géographiques (longitude, latitude et altitude) et les données massives de génomique. Ces avancées permettront de comparer plus un polymorphisme de séquences mais plutôt un polymorphisme de génome…
CHAPITRE I ETUDE DE LA STRUCTURATION GENETIQUE DES POPULATIONS DE LAGOPEDE ALPIN (LAGOPUS MUTA PYRENAICA) DANS LES PYRENEES |