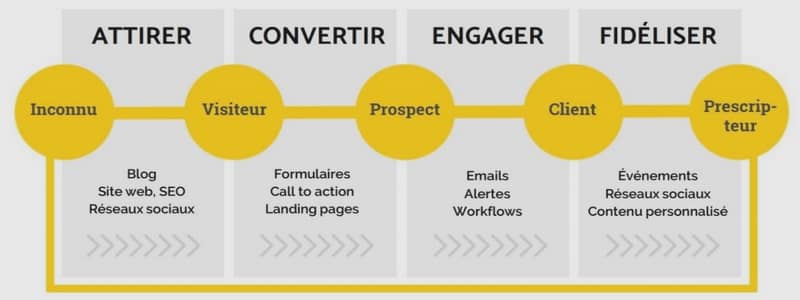
Inhibition de la croissance bactérienne
Dans ce chapitre nous présenterons le concept global appliqué à toutes les expériences afin de déterminer la CMI du céfotaxime sur E.coli. Les expériences ont pour but de quantifier l’effet qu’à la taille de l’inoculum sur la valeur de la CMI. Nous discuterons ensuite de cet effet. Comme décrit dans le chapitre précédent, nous marquons une concentration en antibiotique par une intensité fluorescente. Les fluorophores utilisés sont une dispersion de colloïdes fluorescentes (de 3 m de diamètre). La dilution en continue des collïdes par le LB donne le profil de l’intensité fluorescente initiale du train de goutte (figure 4.1 p.118).Techniquement, nous formons les gouttes dans sept mètres de capillaires, maisles stockons et manipulons sur 3; 5 m, ce qui permet de contenir environ 3000 gouttes. Sachant que nous avons des espaceurs, nous produisons au final environ 1500 gouttes d’intérêt. Nous contrôlons la pente de dilution () pour produire un gradient de concentration dans mille gouttes (sur les 1500). Il est plus sûr de faire cela que de tenter d’avoir le gradient sur toutes les gouttes. En effet, la sélection du train (section 3.4 p.94) s’effectue en fonction du nombre de gouttes qu’on a préalablement choisi. Nous avons pu observé, lorsque nous fermons des vannes pour sélectionner un train, que les gouttes continuent leur écoulement. Ce phé-le gradient de concentration en antibiotique sur un nombre de gouttes inférieur au nombre du gouttes constituant le train.
Inhibition de la croissance bactérienne
nomène est certainement du fait de la déformation des matériaux qui reprennent leurs dimensions d’origines, lorsqu’ils ne sont plus soumis aux pressions crées par les débits imposés. Le « retour à la normale » fait s’écouler les gouttes dans les capillaires, et certaines sortent du capillaire de mesure. La perte est considérée comme faible (moins de 15%). Nous ne pouvons pas savoir quelles gouttes sont perdues. Si nous produisions notre gradient sur toutes les gouttes du train sélectionné, nous ne pourrions pas conserver la totalité du gradient. Dans ce cas, le problème majeur est que nous ne pourrions pas savoir quelles concentrations nous perdons. En pratique, nous formons les gouttes de la première à la seconde branche (figure 3.9 p.95). Initialement la concentration en céfotaxime est constante. Nous déclenchons en même temps la dilution et le comptage des gouttes pour la sélection. Puis nous comptabilisons 1500 gouttes. A cet instant nous avons 1500 gouttes de concentration constante en céfotaxime dans la seconde branche et 1500, qui contiennent le gradient, dans la première. C’est le moment où nous arrêtons le flux d’eau pour ne plus former de gouttes. Nous poussons donc les gouttes et vérifions que nous avons composé un gradient, par observation du niveau de fluorescence. Si nous avions eût le gradient sur les 1500 gouttes, nous n’aurions pas eût de marge d’erreur concernant la sélection du train. Alors que de cette façon nous avons environ 500 gouttes d’assurance (à concentration constante) contre les pertes dues aux erreurs possibles de manipulations et à l’inertie du système. C’est le moyen employé pour avoir le gradient voulu dans le train sélectionné.
C’est pourquoi le profil se décompose en :temps et de la concentration en antibiotique de 10% des échantillons. Nous pou- vons observer que la majorité des bactéries ont atteint la phase stationnaire au bout des cinq heures. Nous notons également que la densité finale est dépendante de la concentration en antibiotique. Le profil de l’intensité de toutes les gouttes après cinq heures d’incubation permet d’observer la variation du signal mesuré (figure 4.4 p.122). Cette figure compare, pour chaque goutte, l’intensité après une période d’incubation au signal initial. Tout d’abord, à cause des champs de vitesse à l’intérieur des gouttes, la densité de colloïdes et bactéries est distribuée en conti- nue dans la goutte. Ces particules fluctuent légèrement au niveau du plan focal de détection. Cela produit donc une légère variation de quantité de fluorophore dans le champs de détection selon les gouttes. De ce fait, nous imposons un filtreEn travaillant donc avec les courbes de lissage, nous observons que les signaux des 700 première gouttes n’ont pas, ou très peu, évolués. Ceci est dû au fait que ces gouttes sont justement celles qui sont soumises à de fortes concentrations en céfotaxime (figure 4.2 p.119). Nous notons d’ailleurs, que ce sont les gouttes qui ne se sont pas multipliées (figure 4.3 p.121). Cela signifie donc que les concentra- tions impliquées ici, inhibent la croissance bactérienne. Lorsque la courbe noire de lissage (figure 4.4 p.122) se trouve en dessous de la courbe jaune, nous avons per- dus en intensité. Ce phénomène est dû à la décroissance du nombre de molécules fluorescentes dans un état excitable (le photoblanchiment) à cause des nombreux passages devant le détecteur.