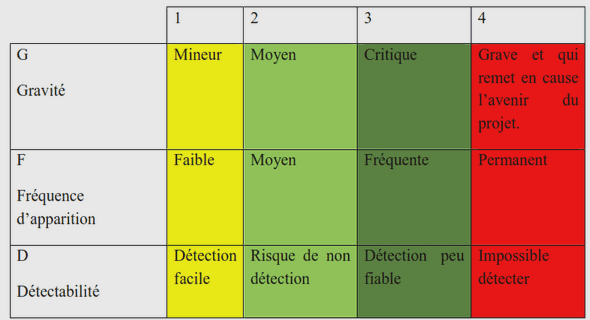
Le Mésolithique un contexte plus favorable au parasitisme
Les chasseurs-cueilleurs du Mésolithique
En Europe de l’Ouest, les débuts du Mésolithique sont classiquement associés avec le début de l’Holocène (9 600 B.C). Cette période a vu les dernières populations de chasseurs-cueilleurs-collecteurs (Rozoy, 1978), derniers représentants d’un mode de vie fondé sur la chasse, la cueillette et la collecte (Thévenin, 1998).
Le Mésolithique ne peut être défini à l’échelle large de l’Europe. Il est plutôt considéré comme une « mosaïque culturelle » (Rozoy, 1978), composée d’ensembles différenciés adaptés à leurs environnements respectifs (Kozlowski, 2009 ; Bailey & Spikins, 2008). Généralement, il n’est pas perçu comme une rupture franche par rapport à la période précédente, le Paléolithique, mais plutôt comme un continuum avec des innovations techniques et sociales adaptées aux changements de biocénoses (Valentin, 2008 ; Valdeyron, 2013). Le Mésolithique a, dans un premier temps, été « mal perçu » par la communauté scientifique.
Une explication vient de la position du Mésolithique, entre le Paléolithique et le Néolithique (Guillomet-Malmasari, 2014), périodes sur lesquelles les chercheurs se sont principalement concentrés . De surcroît, la microlithisation des vestiges lithiques les plus typiques de cette période, les armatures de flèches, a desservi la reconnaissance et la caractérisation de ce technocomplexe jusqu’à la mise en place plus systématique de tamisages. Les années 70 (Kozlowski, 1976 ; Rozoy, 1978) et surtout la fin des années 1980 ont vu la généralisation de l’archéologie préventive qui a entraîné la découverte de gisements bien conservés en plaines alluviales, ou en fond de vallées, ou de fosses ou de structures en creux (Souffi et al., 2007 ; Verjux, 2015). Ces nouvelles découvertes combinées à la reprise de fouilles anciennes en grottes et abris ont octroyé au Mésolithique une place plus significative au sein des problématiques de recherche.
L’industrie lithique de cette période a, dans un premier temps, été essentiellement appréhendée à travers l’étude typologique des armatures. De manière générale, ces éléments caractéristiques ont subi une microlithisation et une standardisation importante, en lien avec la diffusion de l’arc nécessitant l’emploi d’armatures légères (Rozoy, 1978). Il ne faudrait toutefois pas résumer l’industrie lithique mésolithique à ce constat. De fait, certains gisements ont délivré un panel d’outils plus diversifiés et notamment du macro-outillage (Barrière, 1974).
De façon synthétique, pour le Sud de la France, le Mésolithique peut être scindé en deux. La première étape, celle du Sauveterrien, a été caractérisée par le débitage de lamelles étroites et relativement irrégulières par percussion directe à la pierre, et retouchée pour obtenir des microlithes (Angelin, 2017). La seconde, qui a vu le développement du Castelnovien et des cultures qui lui sont contemporaines, s’est distinguée par l’apparition de lames plus normées et plus larges débitées par percussion indirecte ou par pression, accompagnées de trapèzes et de lames à coches (Binder, et al., 2012, Gassin et al., 2013, Marchand & Perrin, 2017).
L’industrie lithique de la fin du Mésolithique de la Péninsule ibérique et du sud de la France a livré des trapèzes réalisés sur de petites lames, puis des pointes triangulaires (Guilaine, 2018). Ces innovations techniques et culturelles se sont répandues assez rapidement à partir du VIIe millénaire selon un axe Est-Ouest et Sud-Nord à partir du Bassin méditerranéen (Perrin et al., 2009 ; Marchand & Perrin, 2017).
Au cours du Mésolithique, l’exploitation des ressources animales et végétales s’est diversifiée à la faveur des nouveaux biotopes régionaux. La cueillette et la collecte auraient occupé une part importante de l’économie de subsistance (Clarke, 1976 ; Mason, 2000 ; Valdeyron, 2013). Le site de Noyen-sur-Seine (77), par exemple a révélé l’importance du domaine végétal dans la vie quotidienne des mésolithiques, au-delà de l’alimentation. Dans le nord de la France, les analyses tracéologiques ont démontré l’importance du travail des matériaux d’origine végétale, rejoignant les observations déjà réalisées en Belgique (Gueret, et al., 2014). De la même manière, les lames à coches du Second Mésolithique ont également été rattachées au travail du végétal sur différents sites français, aussi bien dans le Nord que dans le Sud (Gassin et al., 2013).
En ce qui concerne le domaine cynégétique, un large spectre d’espèces animales a été consommé, avec toutefois des disparités régionales (Marchand, 2012). Tous les milieux ont été exploités. Les populations des façades littorales ont eu une alimentation principalement tournée vers la mer, se traduisant par la consommation de crustacés, d’oiseaux, de mammifères marins et de poissons. À titre d’exemple, à Teviec et Hoëdic, les analyses isotopiques menées sur des restes humains ont montré un ratio de respectivement 50 et 80% de protéines d’origine marine (Marchand, 2012).
À l’intérieur des terres, les grands mammifères comme le sanglier, le cerf, le chevreuil et, de façon plus épisodique, l’aurochs, ont occupé une part importante des cortèges fauniques. Des espèces secondaires ont également été consommées : escargots terrestres, oiseaux, petits mammifères, poissons de rivière. La pêche est attestée à Noyen-sur-Seine grâce à la découverte de plusieurs nasses et d’une pirogue monoxyle (Mordant, 1992).
L’ensemble des données récoltées a eu tendance à illustrer une réduction de la taille des aires de circulation des groupes mésolithiques par rapport aux périodes précédentes, comme semble le montrer l’origine des matières premières lithiques. Certains milieux offrent des ressources consommables tout au long de l’année. À Téviec et Hoëdic l’accumulation des déchets plaide en faveur d’une occupation fréquente et de longue durée avec une grande abondance des ressources océaniques (Dupont et al., 2009 ; Marchand, 2012).
D’autres gisements plus continentaux illustrent ce phénomène. Le « Parc du Château » à Auneau (28) a livré plus d’une soixantaine de structures en creux, dont certaines ont été interprétées comme des fosses de stockage (Leduc & Verjux, 2014 ; Verjux, 2015), attestant de l’occupation régulière et pérenne du site. La fosse n°32, à titre d’exemple, a livré des restes de diverses espèces (aurochs, cerf, sanglier), représentatifs d’une consommation tout au long de l’année (Verjux, 2015). Le Cuzoul de Gramat (46) a également fourni des indices d’une possible occupation longue, notamment à la mauvaise saison, qui se traduirait entre autres par l’investissement, dans l’espace habité, caractérisé par de nombreuses structures de combustion (Valdeyron, 2013) dont une au moins peut renvoyer à un fonctionnement hivernal.
D’autres types de données, notamment paléoenvironnementales, ont mis en évidence des traces d’anthropisation du milieu par des populations mésolithiques. À Noyen-sur-Seine, une augmentation sensible du taux de plantes rudérales et des variations des courbes des milieux riparriens ont été interprétées comme une intervention sur le milieu des populations mésolithiques durant les VIIIe et VIIe millénaire B.C, avec une fréquentation régulière du lieu (Leroyer, et al., 2004).
Le Mésolithique a également été marqué par un accroissement des pratiques funéraires, par rapport à la période précédente. Une trentaine de sites funéraires ont été mis au jour sur le territoire français pour cette période dont dix-neuf datés à partir de restes humains (Meiklejohn et al., 2010). Le nombre de sépultures retrouvées est largement supérieur aux périodes précédentes, mais aucune tendance générale dans les pratiques n’a pu être dégagée si ce n’est un grand éclectisme (Valdeyron, 2013). De fait, une grande variabilité existe dans le domaine funéraire. Des sépultures ont été découvertes isolées, ou implantées au cœur de l’habitat. D’autres ont été regroupées en nécropole (Ghesquière & Marchand, 2010) dans des contextes géographiques et topographiques très différents comprenant des abris sous roche, des cavités, des grottes karstiques, ou encore en plein air.
L’innovation néolithique et son processus de diffusion
La néolithisation, et les transformations socio-économiques fondamentales qu’elle implique, sont un des sujets majeurs des études pluridisciplinaires sur la Préhistoire récente réunissant anthropologie physique/sociale, archéozoologie, archéobotanique, génétique, paléoparasitologie et sciences paléoenvironnementales. Depuis longtemps, le processus de néolithisation est perçu en Europe comme le résultat d’une colonisation des agriculteurs éleveurs venus du Proche-Orient (Childe, 1925). Il a représenté une transformation majeure du mode de vie des populations, constituant en cela une véritable « révolution » (Childe, 1936). Au Proche-Orient, la néolithisation a correspondu à la transformation progressive des sociétés humaines. Celles-ci sont passées de nomades chasseurs-cueilleurs-collecteurs aux sédentaires agropasteurs. Les sociétés de prédation étant progressivement devenues des sociétés de production, les comportements sociaux et symboliques, et les techniques ont été transformés et se sont diffusés progressivement en Europe (Binder, 2000 ; Goring-Morris & Belfer-Cohen, 2011 ; Cauvin, 2013).
Le passage d’un mode de vie de chasseurs-cueilleurs-collecteurs nomades à celui de producteurs sédentaires s’est opéré, lentement, sur près de 4 000 ans au Proche-Orient (Hauptmann, 2011). Dans un premier temps, une stabilisation des populations humaines a eu lieu. Les premières habitations pérennes sont apparues dès 11 000 B.C. Elles étaient de forme circulaires ou semi-circulaires (Belfer-Cohen & Bar-Yosef, 2000 ; Belfer-Cohen & Goring-Morris, 2013). L’utilisation de l’argile dans les constructions est attestée dès 10 000 B.C, la céramique apparaissant quant à elle vers 7 500 B.C (Mazurié de Keroualin, 2003).
Les différents processus de domestication ayant eu lieu au Proche-Orient trouvent leurs origines dans des établissements de petite taille. Ces processus ont amélioré la productivité, la prévisibilité des ressources alimentaires des communautés devenues sédentaires (Zeder, 2017). Les données génétiques ont démontré que quelques lignées d’espèces animales sauvages ont été capturées à des fins de domestication (Zeder, 2017, Daly et al., 2018). L’aurochs (Bos primigenius), la chèvre sauvage (Capra aegagrus), le mouflon (Ovis orientalis) et le sanglier (Sus scrofa) ont été domestiqués à peu près au même moment, vers 8 400 B.C pour le premier et vers 8 500 B.C pour les trois autres (Helmer et al., 2005 ; Zeder, 2011 ; 2012 ; 2017 ; Ottoni et al., 2013). Il s’agit des quatre espèces principales à avoir été domestiquées. Les dernières études effectuées sur la domestication des plantes et des animaux en paléogénétique ont eu pour effet de vieillir les débuts de cette domestication.
Les différences morphologiques entre espèces sauvages et espèces domestiques sont intervenues avec un long décalage (Tanno & Willcox, 2006 ; Vigne 2011). Les plus anciennes dates assurées de domestication végétale11 au Proche-Orient sont de 8 600 B.C à Aswad, 8 500 B.C à Çayönü (Wilcox, 2014). Depuis quelques années, des découvertes telles que des meules ou encore des espaces entièrement dédiés à la mouture suggèrent cependant une agriculture pré-domestique dès 9 500 B.C, notamment à Dhra (9 500 B.C) et à Jerf el Ahmar (9 250 B.C) (Willcox et al., 2008 ; Stordeur, & Willcox, 2016). Les céréales sauvages telles que l’orge, (Hordeum spontaneum), le blé (Triticum boeticum thadouar et Triticum diccocoides) les légumineuses, les lentilles (Lens culinaris), et les pois (Pisum sativum) ont été concernées. Les diagrammes polliniques ont également mis en évidence le développement des plantes adventices (Wilcox, 2014).
Les premières traces d’expansion des sociétés agropastorales hors du Proche-Orient ont été mises au jour à Chypre sur le site de Shillourokambos vers 8 400-8 300 B.C, (Vigne, et al., 2017). Ce nouveau mode de vie s’est ensuite diffusé dans toute l’Europe par l’Anatolie vers 6 800 B.C, selon un modèle arythmique12 . Les populations ont dû alors s’adapter aux différentes composantes des nouveaux écosystèmes qu’elles ont rencontrés13 (Guilaine, 2000 ; 2003 ; 2013; Mazurié de Keroualin, 2003 ; Guilaine & Manen, 2007 ; Bocquet-Appel, 2009 ; Bocquet-Appel, 2009 et al., 2012 ) selon deux voies de néolithisation (Coward et al., 2008), comme l’ont confirmé les études paléogenétiques (Mathieson et al., 2018).
L’origine de la néolithisation pour les régions du nord-ouest de l’Espagne et du sud-ouest de la France, sur lesquelles se concentre notre travail, sont issues du courant proche-oriental. Les sources archéologiques et les analyses génétiques plaident pour une diffusion démique », c’est- à-dire fondée sur l’expansion démographique des populations néolithiques qui ont colonisé de nouveaux territoires avec une incorporation du fonds indigène mésolithique (Poplin et al., 1986 ; Ammerman & Cavalli-Sforza, 1984 ; Renfrew, 1987 ; Vigne, 2004 ; Edwards et al., 2007 ; Perlès, 2010 ; Rigaud, 2011 ; Cauvin, 2013 ; Hofmanová et al., 2016 ; Harris, 2017). Un courant de néolithisation a suivi le cours du Danube, depuis la Hongrie jusqu’à l’Atlantique.
Il est caractérisé par une tradition technique et culturelle de céramiques aux décors à bandes linéaires, par des habitations à charpente massive avec des plans très normalisés et par spectre de faune domestique dominé par les bovidés (Cauwe et al., 2007). Ce courant Rubané ou LBK a atteint la France vers 5 300-5 200 B.C (Foley et al., 2013). L’autre courant, a suivi les côtes méditerranéennes avec des installations en Italie du Sud-est et en Sicile à partir de 6 000 B.C (Binder, 2000 ; Bernabeu Aubán et al., 2017). Il a été caractérisé, en atteignant les côtes françaises et espagnoles par l’Impresso-Cardial. Il est apparu sur les côtes méridionales de la France actuelle à partir de 5 750 B.C (Perrin et al., 2017), puis la navigation côtière l’a porté jusqu’à la péninsule ibérique (Zilhão, 2001 ; Gamba et al., 2012 ; Isern et al., 2017 ; Manen, 2018).
« Il semble en effet que le modèle de la « vague d’avancée » ait été conçu de façon trop générale, sans tenir compte de la variabilité culturelle qui caractérise, d’emblée, les débuts du Néolithique européen. Par variabilité culturelle, on n’entend pas seulement les manifestations identitaires (sitologie, morphologie de l’habitat, productions matérielles et idéologiques) mais aussi les processus d’adaptation aux contextes écologiques et donc les comportements économiques et les « genres de vie ». De fait l’histoire du premier Néolithique en Europe est celle d’une série d’adaptations à des environnements variés auxquelles se surimposent, parallèlement, des renouvellements, des transformations du paysage culturel. Ces mutations, ces métamorphoses ont entraîné périodiquement des recompositions profondes qui ont inévitablement perturbé le rythme des déplacements. » (Guilaine, 2000-2001, p.268)
L’expansion des populations agropastorales le long des côtes méditerranéennes et la colonisation de ce littoral n’a pas été un processus uniforme et continu. L’étude technologique de la céramique a notamment démontré que les populations associées à la première expansion Impressa sur l’aire liguro-provençale, (Arène Candide, Pendimoun) provenaient de deux groupes distincts (Gomart, 2017). La céramique y est caractérisée par un décor à impressions et incisions, comportant de nombreuses variabilités. Ce degré de variabilité dans les assemblages augmente à mesure que les populations se sont déplacées vers l’Ouest, ce que certains auteurs interprètent comme le résultat d’une première vague de colonisation trouvant ses racines dans l’Italie du Sud (Guilaine et al., 2007 ; Zilhão, 2011 ; Manen, 2014 ; Guilaine, 2017).
Concernant le territoire français, les premières implantations ont été retrouvées en Provence (Pendimoun) et en Languedoc (Pont de Roque-Haute). Elles sont perceptibles à travers les assemblages archéologiques et polliniques (Jalut, 1995 ; Puertas, 1999 ; Guilaine, et al., 2007 ; Manen, 2014 ; Guilaine, 2017)..
Un nombre plus important de sites apparaît autour de 5 600 et 5 400 B.C (Manen, 2014) avec la mise en place du Cardial. Le Cardial tyrrhénien d’une part, retrouvé en Italie dans le Latium et en Toscane, en Corse et en Sardaigne. Et d’autre part, le Cardial franco/ibérique sur le pourtour méditerranéen occidentale (Manen, 2014), identifié en Languedoc et en Provence vers 5 500-5 400 B.C pour le territoire Français. Pour la péninsule Ibérique, il est identifié en Aragon, en Catalogne, dans la province de Valence et en Andalousie, où les premiers gisements sont datés autour de 5 600 B.C et vers 5 500 B.C au Portugal (Bernabeu Aubán et al., 2017).
Le Cardial franco/ibérique a couvert tout le pourtour de la Méditerranée occidentale (Manen, 2014), avec des sites de plein air, des grottes et des abris sous roche. Les populations y pratiquaient un système agricole mixte avec la présence de blé et d’orge, combiné avec des activités pastorales basées préférentiellement sur les ovins/caprins, concernant les ressources carnées (Helmer & Vigne, 2004).
14 « The neolithisation experiences are the result not of a narrowly typed culture but rather of distinct uncoordinated pioneer actions that are manifested in distinct thematic variety which differs from site to site according to the navigations and the places selected for the foundation of small “farmstead” settlements. » [Les expériences de néolithisation ne sont pas le résultat d’une culture étroitement typée mais plutôt d’actions pionnières distinctes et non coordonnées qui se manifestent dans une variété thématique distincte qui diffère d’un site à l’autre selon les navigations et les lieux choisis pour la fondation de petits etablissement “fermes”.] (Guilaine, 2017, p.8)
Les industries lithiques du cardial au sens large reposent sur la coexistence de deux chaînes opératoires distinctes (Binder, 1987; Briois, 2005; Perrin, 2014) :
• Une production laminaire réalisée, sur des matières premières locales ou régionales. Les support ont été obtenus par percussion indirecte, ils ont ensuite été utilisés bruts ou retouchés principalement pour fabriquer des armatures de pointe de flèche.
• Une production d’éclats, plus expédiantes, réalisé aux dépens de ressources locales. Ces manières de faire les procédés techniques qui sous-tendent et les types d’outils fabriqués s’inscrivent nettement en rupture avec les industries du Mésolithique (Binder, 1987).
Des interactions et des hommes
La genèse et la dispersion des premières sociétés agropastorales ont entraîné des changements dans l’équilibre des écosystèmes. Les populations humaines néolithiques ont en effet exercé de nouvelles pressions sur leur environnement à travers entre autres, la domestication des animaux et des plantes. Ces changements globaux ont eu un impact sur la biodiversité, impliquant des perturbations dans la transmission des agents pathogènes, que ce soit des micros ou des macros parasites (Cockburn, 1971 ; Omran, 1971 ; McNeil, 1976 ; Barret et al., 1998 ; Morand, et al., 2014).
Des changements d’hôtes peuvent s’opérer si l’Homme procure au parasite un environnement similaire à celui de son hôte naturel, ou un nouvel hôte (Bowden & Drake, 2013). Connaître l’apparition des agents infectieux, leurs aires de répartition et leurs extinctions est un préalable fondamental pour appréhender les risques infectieux et s’interroger sur le rôle de régulation qu’ils ont pu jouer dans le développement biologique et démographique des populations humaines passées (Mitchell, 2013).
L’existence d’agents pathogènes implique une notion de transmission. Afin d’assurer leur propagation, les agents parasitaires ont élaboré différents moyens de transmission pour entrer en contact avec leurs hôtes et effectuer leurs cycles.
Transmission par consommation
Les helminthes retrouvés dans nos sédiments archéologiques possèdent tous ce moyen de transmission, qu’ils soient infestants pour les animaux ou pour l’humain. Il s’agit majoritairement de mésoparasites. Ils se situent dans le tractus digestif de l’hôte, en communication avec le milieu extérieur, de sorte que les propagules sont libérées par les voies naturelles. Ils s’insèrent dans la chaîne alimentaire via la consommation de végétaux ou de viandes. Pour les végétaux, on peut citer l’exemple de la douve du foie (Fasciola hepatica) dont les stades infestants (métacercaires) se collent sur les feuilles de végétaux et sont broutés par les ruminants ou ingérés par l’Homme. Pour la viande, ce sont par exemple les larves infestantes du Taenia sp., ou « vers solitaire », qui se situent dans les muscles du porc (Taenia solium) ou de bœuf (Taenia saginata).
Les œufs peuvent également être associés à l’alimentation sans pour autant être dépendants d’une consommation bien précise comme pour le Trichuris trichiura, parasite de l’Homme dont des œufs ont été retrouvés dans la momie Ötzi (Aspöck et al., 1996 ; 2000). Ces œufs s’embryonnent directement dans le milieu extérieur. L’infestation se fait par ingestion accidentelle d’œufs embryonnés à travers les aliments souillés, des mains sales porteuses de terre ou l’eau de boisson. Ce type de parasitose est dépendant du degré d’hygiène.
Transmission par contact
Chez les hôtes, la vie sociale et la densité de la population ont un impact direct sur la transmission des vecteurs parasitaires. Dans des conditions de promiscuité extrême, des niveaux élevés de parasitisme peuvent ce produire. À titre d’exemple, les épidémies de gale coïncident avec toutes sortes d’accroissements de promiscuité : guerres, rassemblements hippies et concerts de rock, de même que pour les poux (Pediculus capitis) (Combes, 1995). Les maladies et les parasites sexuellement transmissibles telles que le VIH ou les morpions (Phtirius inguinalis) se transmettent également directement d’hôte à hôte (Combes et al., 2018).
Transmission par stade libre infestant
Le mode de transmission par stade libre est inféodé à des contextes aquatiques ou des boues. Pour l’Homme, le nématode Ancylostoma duodenale a ce moyen de transmission. Ses larves pénètrent à travers la peau, comme chez les Schistosomes, dont les cercaires (larves) infestent l’hôte définitif par voie transcutanée, occasionnant des lésions des tissus.