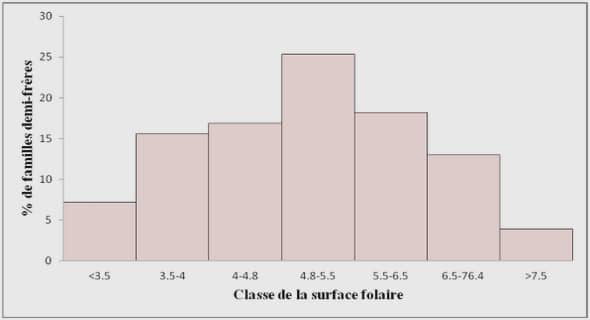
Les facteurs contrôlant la répartition des espèces sont des connaissances essentielles pour comprendre la dynamique des écosystèmes forestiers. Ces connaissances peuvent permettre, entre autre, de prévoir la réponse des écosystèmes ou des arbres aux changements climatiques et de mieux aménager le territoire de façon durable. En effet, un réchauffement climatique sans précédent en terme de vitesse et de magnitude est anticipé (GIEC 2007). Ce réchauffement pourrait avoir une importante répercussion sur la dynamique des écosystèmes. En effet, le climat peut influencer la physiologie, la phénologie (Kullman, 2007; Delbart et al., 2008) et la croissance (Huang et al., 2007; St-Germain et Krause, 2008) d’une espèce. Plusieurs études ont montré la migration ou l’extinction locale d’espèces en raison de changement climatique (Lloyd et Fastie, 2003; Parmesan, 2006; Kullman, 2007). Ainsi, le développement de modèle prédictif de la migration des espèces est très important afin de minimiser les conséquences potentiellement négatives des changements climatiques sur les écosystèmes forestiers.
Les modèles prédictifs actuels se basent principalement sur la niche réalisée des espèces (niche-based models) pour faire leur prédiction (Ressources naturelles Canada 2011). Ces modèles sont ainsi basés sur la présence ou l’absence d’espèces à certaines températures et supposent qu’avec le réchauffement climatique, ces espèces migreront pour s’établir à ces mêmes températures. Ces modèles ne tiennent pas compte du temps de réaction de l’espèce et de sa possibilité de dispersion. D’autres modèles tentent d’intégrer des données de type phénologique (process-based models) afin d’avoir un portrait plus réaliste pour chacune des espèces (Morin et al., 2008).
La répartition d’une espèce est la résultante d’une combinaison complexe de facteurs. Certaines études s’accordent pour dire que le climat est un élément prépondérant dans la répartition des espèces à une grande échelle (Prentice et al., 1992; Wang et al., 2006). Toutefois, il semble que la limite nordique de la répartition des arbres ne soit pas uniquement contrôlée par le climat. Les perturbations naturelles, les conditions édaphiques et la compétition interspécifique peuvent aussi jouer un rôle important à une échelle plus régionale ou locale (Woodward, 1987; Richardson et Bond, 1991). L’importance de ces facteurs dépendraient de chacune des espèces. Ainsi, afm de prévoir correctement la réaction des espèces et de leur écosystème, il est essentiel d’étudier davantage les facteurs qui influencent la répartition d’une espèce pour que ces résultats soient intégrés dans les modèles de prédiction.
Peu d’études se sont penchées simultanément sur plusieurs facteurs explicatifs de la limite de répartition, préférant se concentrer sur une seule, généralement le climat. De plus, au Québec, aucune étude ne s’est réellement penchée sur l’étude d’une gamme de facteurs potentiels limitant la répartition du thuya de l’Est (Thuja occidentalis L.). Certaines études ont émis l’hypothèse d’une fréquence des feux trop grande (Heinselman, 1973) ou d’un climat peu propice (Tardif et Stevenson, 2001), mais l’importance relative de ces facteurs et de leurs interactions sur la répartition nordique du thuya de l’Est est méconnue.
L’effet de facteurs climatiques sur la répartition d’une espèce
Le climat a souvent été suggéré comme étant un facteur contrôlant la répartition nordique des espèces (Richard, 1993; Carcaillet et al., 2001). En effet, à la limite nordique de leur répartition, certaines espèces ont des populations de petite taille, une faible croissance et un faible taux de reproduction, conditions qui peuvent être attribuées au climat (Pigott et Huntley, 1981; Payette et Delwaide, 1994; Kullman, 2007). D’autre part, certaines études ont montré que la limite de répartition de certaines espèces d’arbres, notamment le sapin baumier (Abies balsamea L.) et thuya de l’Est, était située plus au nord lorsque les températures étaient plus chaudes, ce qui suggère un effet du climat sur la répartition (Richard, 1993; Ali et al., 2008).
Impact du climat sur la survie des arbres
Le climat peut affecter directement la survie des arbres en infligeant des blessures dues au froid, au feuillage et aux branches, ou par un nombre insuffisant de jours de croissance ou de précipitations (Woodward, 1987; Kullman, 1998). Sakai et Weiser (1973) ont d’ailleurs étudié l’effet de très basses températures sur la survie des branches et bourgeons. Ils ont démontré que plusieurs espèces boréales pouvaient tolérer des températures aussi froides que -80°C. Il est néanmoins suggéré que l’effet du dessèchement hivernal pourrait avoir un impact plus important que le climat sur la survie des arbres (Hadley et Smith, 1983). Les variations de température et de précipitation peuvent également influencer le taux de croissance radiale d’une espèce pour une même latitude (Kozlowski et al., 1991; Tardif et Bergeron, 1997; Tardif et Stevenson, 2001; Heinrichs et al., 2007; Vila et al., 2008) ou le long d’un gradient latitudinal (Pam, 2003; St-Germain et Krause, 2008; Huang et al., 2010).
Impact du climat sur le potentiel reproductif
L’expansion de l’aire de répartition d’une population d’arbres est reliée à sa régénération. Selon Caspersen et Saprunoff (2005), la régénération d’une espèce dépend de la production de graines viables et de sites adéquats pour la germination. Toutefois, les conditions ambiantes doivent également être propices à la survie des semis. Les différentes étapes de la régénération peuvent être affectées par certains facteurs et mettre en péril la survie de l’espèce. Plusieurs études traitent de l’effet du climat à la limite nordique sur le processus reproductif des arbres en comparant la régénération à différentes latitudes (Pigott et Huntley, 1981; Flannigan et Woodward, 1993; Houle et Filion, 1993; Tremblay et al., 2002; Messaoud et al., 2007a). Ainsi, à la limite de répartition des arbres, le climat peut empêcher une forme de croissance érigée et la production de graines viables, essentielles pour la dispersion (Asselin et Payette, 2006). De plus, chez les peuplements d’érable rouge (Acer rubrum L.) situés plus au nord, la production de samares est limitée possiblement en raison des épisodes de gel printanier, qui peut grandement diminuer le nombre de fleurs et de graines fertilisées (Tremblay et al., 2002). Pour leur part, Pigott et Huntley (1981) ont montré que le tilleul à petites feuilles (Tilia cordata Mill.) ne pouvait se reproduire de façon sexuée à sa limite nordique. Ils se sont penchés spécifiquement sur les différentes phases de production d’une graine et ont démontré que le climat rigoureux limite l’allongement du tube pollinique et l’atteinte de l’ovule.
Le succès de germination des graines et la survie des semis sont aussi affectés par le climat. Pour certaines essences, dont le frêne blanc (Fra.xinus americana L.), l’humidité constante est une variable des plus importantes pour la germination et la survie des semis (Simard et al., 1998; Cornett et al., 2000b; Messaoud et Houle, 2006). Ensuite, la température de l’air, par exemple le nombre de degrés-jours de croissance et le nombre de jour de gel printanier, semblent aussi jouer un rôle dans l’établissement des semis (Sirois, 2000; Tremblay et al., 2002; Asselin et al., 2003; Kullman, 2007). Les graines de pin rouge (Pinus resinosa Ait.), pour leur part, requièrent des températures de plus de 1 Ü°C pour germer (Flannigan et Woodward, 1993). De même, les semis de sapin baumier semblent être sensibles au climat puisque la densité de semis diminue au nord et ce même si la densité de sapins matures dans un peuplement y était plus élevée (Messaoud et al., 2007a).
La reproduction sexuée peut être remplacée par un mode asexué en raison de contraintes environnementales. La reproduction végétative peut permettre à certaines espèces de se maintenir lors d’une perturbation ou d’une interruption dans leur cycle de reproduction. Le cycle reproductif peut être altéré notamment par une plus petite saison de croissance, par des températures froides ou par la fertilité du sol (Staneck, 1961). L ‘épinette noire (Picea mariana Mill.) et l’érable rouge se propagent majoritairement par reproduction asexuée à la limite nordique de leur répartition pour contrer le climat rigoureux (Fayette, 1979; Tremblay et al., 2002; Lloyd, 2005). Les peuplements d’épinette noire peuvent être composés de plus de 50% de clones à la limite des arbres, possiblement en raison du climat rigoureux qui empêche la germination des graines (Fayette, 1979; Lloyd, 2005).
L’effet des facteurs édaphiques et topographiques sur la répartition d’une espèce
La répartition d’une espèce peut être influencée par les conditions édaphiques environnantes. Une hypothèse soulevée pour expliquer la limite de répartition d’une espèce est la raréfaction d’habitats favorables qui peut être due à un changement dans les types de dépôt ou de drainage. Par exemple, Meilleur et al. (1997) ont démontré le manque de sites propices au pin rigide (Pinus rigida Mill.) à sa limite nordique dans le sud du Québec, ce qui l’empêcherait d’agrandir sa répartition plus au nord. Toutefois, les facteurs édaphiques expliquent rarement complètement la limite de répartition d’une espèce mais font plutôt partie d’une équation de plusieurs facteurs. Mes saoud et al. (2007b) ont montré que la répartition des peuplements de sapin baumier était limitée par une diminution de sites potentiels favorables dans la pessière à mousse. Toutefois, le sapin baumier y était incapable de coloniser tous les sites favorables. Ces résultats suggèrent donc que la diminution de l’abondance des sites ne peut expliquer complètement la répartition du sapin. En Alaska, la présence de l’épinette noire est expliquée à la fois par la fréquence des feux et le pH des sols (Lloyd et al., 2007). De plus, Lalonde (1991) a montré que la répartition de l’érable rouge était influencée par le climat mais également par le paysage. À la limite de son aire de répartition, l’érable rouge se trouve à de plus hautes altitudes, où il est confmé au versant sud-est. L’altitude lui procure un avantage thermique et l’orientation sud augmente aussi la quantité de radiations en plus de le protéger des vents de l’ouest et des feux qui sont dirigés par le vent (MacHattie, 1961). Sa limite de répartition coïncide également avec une raréfaction des collines ce qui peut limiter davantage sa distribution.
L’effet des perturbations naturelles sur la répartition d’une espèce
La forêt boréale de l’Ouest québécois est grandement perturbée par les feux. Les feux y sont très fréquents et peuvent survenir en moyenne tous les 135 ans (Bergeron et al., 2004). Les feux de la forêt boréale sont une perturbation majeure qui influence la composition et possiblement la répartition des espèces (Rowe, 1973; Suffiing, 1995; Bergeron, 2000). Payette et Gagnon (1985) ont constaté que les régimes de feu en combinaison avec le climat contrôlaient la limite des arbres. En effet, certaines espèces requièrent le passage du feu pour se maintenir dans le paysage alors que pour d’autres, la répartition est restreinte par la fréquence et l’intensité des feux. Desponts et Payette (1992) ont montré que le pin gris (Pinus banksiana Lamb.) dépend d’un cycle de feu à courts intervalles pour maintenir ses populations à sa limite nordique puisqu’une chaleur intense est essentielle pour ouvrir les cônes et relâcher les graines. Pour sa part, la répartition du pin rouge semble limitée par les feux forestiers puisqu’il semble posséder une dépendance aux feux de moyenne ou faible intensité. De tels feux permettent de supprimer les espèces compétitrices en sous-couvert et de créer des ouvertures dans la canopée pour sa régénération. Toutefois, les feux de grande intensité, que l’on trouve majoritairement au nord de sa répartition, semblent restreindre l’établissement plus au nord de cette espèce (Bergeron et Gagnon, 1987; Flannigan, 1993; Flannigan et Bergeron, 1998). La présence du genévrier commun (Juniperus communis L.) semble aussi être entretenue par des feux de moyenne intensité sur les îles du Lac Duparquet, ce qui apparaît comme un facteur important dans sa répartition (Diotte et Bergeron, 1989). D ‘autres études démontrent également l ‘influence négative du feu dans l’aire de répartition de certaines espèces. Ils ont montré qu’alors que les feux étaient moins fréquents pendant certaines périodes de 1 ‘Holocène, certaines espèces sensibles aux feux en ont profité pour agrandir leur répartition plus au nord (Carcaillet et al., 2001; Messaoud et al., 2007b; Ali et al., 2008).
INTRODUCTION GÉNÉRALE |