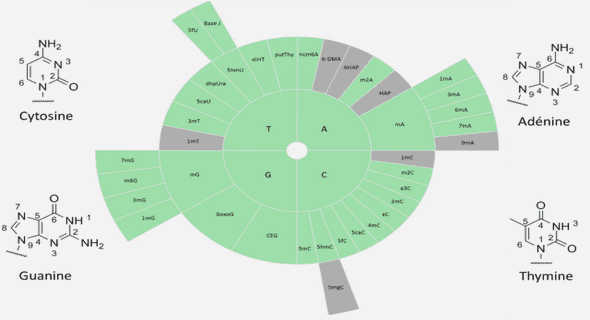
Effets de l’exercice physique
Lors de ce travail de thèse, nous avons également voulu, en dernier lieu, appréhender les effets de l’exercice sur la microcirculation du tissu adipeux chez la souris normopondérée. Nous avons cherché à savoir si, dans un environnement tissulaire différent du contexte pathologique retrouvé chez l’obèse, l’exercice pouvait également constituer un stimulus proangiogénique, capable d’induire un processus de croissance vasculaire au sein des tissus adipeux blancs et bruns. En condition physiologique, la balance angio-adaptative est dans un état d’équilibre dynamique permettant d’assurer la maintenance d’un réseau capillaire structuré, en adéquation avec les besoins métaboliques et en oxygène du tissu adipeux. Si l’exercice physique est en mesure de modifier l’homéostasie vasculaire du tissu adipeux chez la souris saine, par quels mécanismes moléculaires cela peut-il être expliqué? Et quelles conséquences cela peut-il avoir sur les fonctions endocrines et métaboliques du tissu adipeux ? Pour répondre à toutes ces interrogations, nous avons utilisé des souris soumises à régime alimentaire standard et à un protocole d’exercice physique volontaire de 7 semaines, identique au protocole utilisé chez les souris HFS. À la fin des 7 semaines, nous avons évalué les effets de l’exercice sur : 1) la masse et la taille adipocytaire des tissus adipeux blancs épididymal et sous-cutané, ainsi que du brun interscapulaire ; 2) la microcirculation du tissu adipeux estimée via un immuno-marquage au CD31 sur des coupes de tissu adipeux épididymal (EWAT), sous-cutané (ScWAT) et brun (BAT) ; 3) l’équilibre de la balance angio-adaptative en mesurant l’expression du niveau messager et protéique des facteurs angiogéniques VEGFA et TSP-1 dans ces tissus adipeux ; 4) l’implication des protéines Mdm2 et FoxOs dans la régulation de cette balance angio-adaptative ; 5) l’expression des adipokines, des gènes de l’adipogénèse et de la thermogenèse dans les différents tissus adipeux.
Expression de Mdm2, FoxOs et des facteurs angiogéniques dans les tissus adipeux de la souris après exercice volontaire. Les protéines et les ARN messagers ont été extraits des tissus adipeux viscéral (EWAT), sous-cutané (ScWAT) et brun (BAT) provenant des souris contrôles sédentaires (NC) et entrainées 7 semaines (NC ex.). Les niveaux protéiques de Mdm2, FoxO1, VEGF-A et TSP-1 dans l’EWAT (A) et le ScWAT (C) ont été analysés par Western Blot. Les blots ont été lavés et réincubés pour l’α/β tubuline, utilisée comme protéine de référence (NC: n=5 ; NC ex. : n=3). Les niveaux messagers de Mdm2, FoxO1, FoxO3α, VEGF-A et TSP-1 dans l’EWAT (B), le ScWAT (D) et le BAT (E) ont été analysés par PCR quantitative en temps réel. Les valeurs ont été normalisées avec l’ARNm de la protéine ribosomale 18S et exprimées en ratio d’expression des souris contrôles sédentaires (NC)(n=5 par groupe). Données exprimées en moyenne ± SEM. * P<0.05, *** P<0.005 (relatif aux NC). Test-T de Student non-apparié.
Effets de l’exercice physique volontaire sur l’hypoxie, les adipokines et l’activité métabolique des tissus adipeux de la souris. Après marquage immuno-histochimique à l’anticorps HIF-1α sur coupe de tissu adipeux, l’aire de marquage à HIF-1α a été évaluée dans l’EWAT, le ScWAT et le BAT des souris contrôles sédentaires (NC) et entrainées 7 semaines (NC ex.) (A). Les ARN messagers (ARNm) ont été extraits des tissus adipeux viscéral (EWAT), sous-cutané (ScWAT) et brun (BAT) provenant des souris NC et NC ex.. Les niveaux messagers de la leptine, de l’adiponectine, du TNF, de l’IL-6, de PPAR, d’UCP-1, PGC1, Sirt3, cidea dans l’EWAT (B), le ScWAT (C) et le BAT (D) ont été analysés par PCR quantitative en temps réel. (E) Images représentatives de tissu adipeux sous-cutané de souris contrôles sédentaires (NC) et entrainée (NC ex.).
Les résultats de cette étude montrent que les souris entrainées ne présentent pas de perte de poids après 7 semaines d’exercice volontaire (Figure 35A). Elles réduisent en revanche leur pourcentage de masse grasse (-26% ; Figure 35B). La réduction du pourcentage de masse grasse s’explique par une réduction significative de la masse des tissus adipeux blancs viscéral épididymal (-52%) et sous-cutané (-44%), alors que la masse de tissu adipeux brun est augmentée (+65% ; Figure 35C). Au niveau cellulaire, la réduction de la masse adipeuse chez les souris entrainées ne se traduit pas dans le tissu adipeux épididymal par une réduction de la taille moyenne des adipocytes, alors que dans le sous-cutané cette taille moyenne est significativement diminuée de 40% (Figure 35D). Dans le tissu adipeux brun, la taille des adipocytes n’a pu être évaluée de par l’architecture très étriquée du tissu. Mais la présence des gouttelettes lipidiques de grande taille dans les adipocytes bruns est réduite (-61%, non significatif) chez les souris entrainées en comparaison des souris saines sédentaires (Figure 35E). L’évaluation du réseau capillaire dans les tissus adipeux grâce au marquage au CD31 a permis de mettre en évidence une densité capillaire augmentée de 76% dans le tissu adipeux épididymal et de 69% dans le tissu sous-cutané des souris entrainées (Figure 35F). Dans ces mêmes tissus, le ratio capillaire par adipocytes est également significativement augmenté de respectivement 37% et 14% (Figure 35G). Dans le tissu adipeux brun, les 7 semaines d’exercice volontaire ont légèrement augmenté (+16%) l’aire marquée au CD31 chez les souris entrainées (Figure 35H).
La mesure des facteurs angiogéniques a été réalisée dans l’ensemble des tissus adipeux. Elle met en lumière une augmentation significative du ratio VEGF-A/TSP-1 dans le tissu adipeux viscéral épididymal (+172% au niveau protéique et +90% au niveau messager ; Figure 36A et B), le tissu adipeux sous-cutané (+110% au niveau protéique et 43% au niveau messager ; Figure 36C et D) et le tissu adipeux brun (+42% au niveau messager ; Figure 36E) des souris entrainées. Si l’expression de FoxOs n’est pas affectée par les 7 semaines d’exercice volontaire dans le tissu adipeux sous-cutané (Figure 36C et D) et brun interscapulaire (Figure36E), celle de FoxO1 est augmentée au niveau messager (+75% ; Figure 36B) et diminuée au niveau protéique (-32% ; Figure 36A) dans le tissu adipeux épididymal. En revanche, il est observé une augmentation de l’expression du messager et de la protéine de Mdm2 dans l’ensemble des tissus adipeux (de respectivement 50% et 73% dans l’EWAT ; 33% et 120% dans le ScWAT ; +40% du messager dans le BAT).