GENERALITES
Généralités sur les chauves-souris
Bio-écologie et taxonomie
Les chauves-souris ou Chiroptera (du grec « chiros » : main et « ptera » : aile) sont les seuls mammifères doués du vol actif grâce à l’allongement de leurs métacarpes et de leurs doigts (à l’exception du pouce), qui sont reliés entre eux par le patagium pour former les ailes (Neuweiler, 2000). Elles représentent environ un cinquième des espèces de mammifères terrestres dans le monde et sont reparties sur les principaux continents avec environ 1120 espèces (Simmons, 2005), mais de récents travaux de recherches ont décrits de nombreuses autres nouvelles espèces pour la Science.
Ces animaux ont des mœurs nocturnes, ils disposent majoritairement du système d’écholocation pour s’orienter dans le noir et pour localiser et capturer leurs proies. Différents types d’habitats peuvent être fréquentés par les chauves-souris comme gîte diurne à savoir les grottes, les crevasses des rochers, les feuillages ou les creux des troncs d’arbres ainsi que les infrastructures humaines, où elles vivent souvent en colonie.
Les chauves-souris figurent parmi les mammifères les plus lents à se reproduire relativement à leur taille. La majorité des espèces n’engendrent qu’un unique petit pendant la reproduction, et pour certaines espèces la reproduction ne débute que vers l’âge de deux ou trois ans. Pour beaucoup d’espèces, le jeune est mis bas au moment où la nourriture est la plus abondante. Ceci est une stratégie de reproduction, permettant à la mère d’avoir suffisamment de nutriment pour la production de lait, et aux jeunes de survivre pendant au moins les premiers moments de leur vie partiellement indépendants de leurs mères (Racey, 1982).
Les chiroptères constituaient auparavant deux sous-ordres, les Megachiroptera et les Microchiroptera (Dobson, 1875; Peterson et al., 1995). Les Megachiroptera, représentés par la seule famille des Pteropodidae, englobent les chauves-souris de grande taille, à sens visuel développé, et essentiellement à régime alimentaire frugivores et nectarivores (Singaravelan & Marimuthu, 2004). Les Microchiroptera sont généralement de petite taille et réunissent plusieurs familles. Leur sens visuel est moins développé que celui des Megachiroptera mais leur écholocation est plus performante.
Leur régime alimentaire est plus diversifié et est dominé par les insectes. Cependant, des données génétiques ont permis d’adopter une nouvelle classification des chauves-souris, qui regroupent actuellement deux sous-ordres, les Yinpterochiroptera constitués des Pteropodidae et des Hipposideridae (anciennement placé parmi les Microchiroptera), et les Yangochiroptera composés des autres Microchiroptera (Teeling et al., 2002, 2005).
Chauves-souris de Madagascar
Madagascar possède une richesse spécifique importante en faune chiroptérologique, avec 46 espèces (Annexe 1) et un taux d’endémisme de 75 % (Goodman, 2011; Goodman et al., 2011, 2012a, 2012b, 2015a, 2015b, 2016), qui est certainement le niveau d’endémisme le plus élevé chez les chauves-souris dans le monde pour une île tropicale. Cette diversité de chiroptères à Madagascar se distribue en neuf familles : les Pteropodidae, les Hipposideridae, les Emballonuridae, les Nycteridae, les Myzopodidae, les Molossidae, les Vespertilionidae, les Miniopteridae (Goodman, 2011) et les Rhinonycteridae (Foley et al., 2015). La famille des Pteropodidae est composée de trois espèces toutes endémiques de l’île : Pteropus rufus, la plus grande en taille, Eidolon dupreanum, de taille moyenne, et Rousettus madagascariensis, la pluspetite des trois espèces. La famille des Hipposideridae compte deux espèces endémiques de Madagascar, Macronycteris commersoni et M. cryptovalorona, cette dernière étant une nouvelle espèce décrite (Goodman et al., 2016). Ces espèces étaientauparavant placées dans le genre Hipposideros mais suite à une récente révisiontaxonomique, elles appartiennent actuellement au genre Macronycteris (Foley et al., 2017). La famille des Nycteridae est représentée par un genre monospécifique également endémique, Nycteris madagascariensis. Myzopodidae est une famille endémique représentée par deux espèces : Myzopoda aurita et M. schielmanni. La famille des Rhinonycteridae réunit les espèces Paratriaenops furculus, P. auritus et Triaenops menamena, qui étaient auparavant classées dans la famille des Hipposideridae. Les familles des Emballonuridae (4 espèces), Molossidae (8 espèces), Vespertilionidae (11 espèces) et Miniopteridae (12 espèces) sont constituées d’espèces endémiques ou à plus large distribution (Afrique continentale, îles de l’Océan Indien et Moyen Orient).
La diversité spécifique dans la partie Ouest de Madagascar (région sèche) est plus importante que dans la partie Est (région humide), en cohérence avec une plus grande diversité d’habitats utilisables comme gîtes diurnes (notamment d’abondantes crevasses et de grottes) dans la partie occidentale de l’île (Goodman et al., 2005).
Généralités sur les parasites
Nature du parasitisme
Le terme parasite, qui vient du mot grec « parasitos » (« para » : à côté de, « sitos » : aliments), signifie « celui qui réside à la charge d’autrui, qui mange à la table d’un autre » (Harper, 2001). Le parasitisme est alors une forme d’interaction interspécifique où le parasite tire profit d’un autre organisme, appelé hôte, essentiellement en termes de nutrition. C’est un mode de vie très répandu dans le monde du vivant, la plupart des organismes libres n’échappant pas au parasitisme.
Le système hôte-parasite est similaire au système proie-prédateur étant donné qu’il s’agit d’une interaction interspécifique qui privilégie le consommateur en termes de transfert d’énergie. La différence réside dans le fait que les prédateurs éliminent directement des individus dans la population proie par opposition aux parasites qui maintiennent généralement une interaction durable avec leurs hôtes (consommateurs insidieux). Ainsi d’un point de vue évolutif, le parasitisme est le produit de la coévolution entre l’hôte et le parasite, ce qui rend ainsi les parasites théoriquement indissociables de leurs hôtes.
Les parasites tirent de nombreux avantages de la transition d’un mode de vie libre à un style de vie parasitaire. Entre autres, ils bénéficient d’un environnement stable créé par le corps de leurs hôtes, puisque les organismes vivants possèdent des mécanismes de régulation pour maintenir leurs fonctions biologiques dans des conditions favorables (homéostasie). De plus, l’association à des hôtes mobiles leur offre une grande capacité de dispersion. En outre, pour réussir à utiliser les ressources de leurs hôtes, les parasites ont essentiellement restreint leurs propres fonctions aux fonctions nutritives. En effet, chez certains parasites, les fonctions telles que la digestion et la locomotion sont presque entièrement laissées à l’hôte (Marshall, 1981a; Combes, 2001; Lehane, 2005).
Cependant, le passage à un mode de vie parasitaire implique un ensemble de défis adaptatifs (Poulin, 2007). Au cours de l’évolution, les parasites doivent développer des stratégies de dispersion entre les hôtes, qui sont généralement distribués de manière discontinue dans le temps et dans l’espace (Marshall, 1981a; Combes, 2001; Poulin, 2007). Cette contrainte exige des ajustements remarquables dans la biologie des parasites. Ces adaptations vont favoriser les parasites qui ont trouvé des moyens efficaces de dispersion et de localisation de leurs hôtes, et qui en contrepartie ont développé des stratégies de reproduction adéquates permettant de compenser les coûts engendrés par la localisation des hôtes (Marshall, 1981a). De plus, les individus hôtes abritent souvent plusieurs espèces de parasites exploitant les mêmes ressources, les parasites sont alors susceptibles d’interagir entre eux et trouver un moyen de coexister ou d’exclure les espèces concurrentes. De manière générale, étant donné que les parasites dépendent de leurs hôtes pour se nourrir, se reproduire et disperser, les adaptations permettant d’exploiter au maximum leurs hôtes tout en les gardant en vie sont sélectionnées (Combes, 2001).
Différents groupes de parasites
Deux grands groupes de parasites sont définis selon la taille, le mode de reproduction et le type d’infection : les microparasites (virus, bactéries, protozoaires) et les macroparasites (helminthes, arthropodes, et certains métazoaires) (Anderson & May, 1979). En général, les microparasites sont caractérisés par une taille microscopique, une reproduction/division s’effectuant au sein de l’organisme hôte, des temps de génération courts et une durée d’infection relativement courte par rapport à la durée de vie de l’hôte. Par contre les macroparasites sont visibles à l’œil nu, ils ont généralement un temps de génération plus long, leur multiplication directe dans l’hôte est peu fréquente, et leur infestation tend à être persistante durant la vie de l’hôte pendant laquelle ce dernier est continuellement réinfesté.
Les parasites sont séparés en trois grandes classes selon leur tropisme chez l’hôte : les endoparasites, les mésoparasites et les ectoparasites (Morand & Deter, 2007).
Les endoparasites et les mésoparasites sont des parasites internes, c’est-à-dire vivant à l’intérieur de l’organisme hôte. Les endoparasites vivent dans les tissus et les liquides internes (vaisseaux sanguins, vaisseaux lymphatiques) voire à l’intérieur des cellules en tant qu’organismes intracellulaires, tandis que les mésoparasites fréquentent les cavités internes de l’hôte (estomac, poumons, intestins, etc.). Autrement dit, les mésoparasites sont en contact avec les milieux extérieurs au sein de l’hôte. Les ectoparasites sont des parasites externes, c’est-à-dire vivant à la surface de l’hôte.
Par ailleurs, le temps dépensé sur ou dans l’hôte par les parasites permet de catégoriser les parasites en deux groupes majeurs : les parasites temporaires et les parasites stationnaires (Olsen, 1974). Les parasites temporaires ne visitent l’hôte que pour se nourrir, tandis que les parasites stationnaires passent une période définie de leur développement sur ou dans le corps de l’hôte. Ce dernier groupe peut encore être subdivisé en deux sous-groupes selon la durée du temps dépensé sur l’hôte, les parasites périodiques qui vivent sur l’hôte seulement pendant une partie de leur cycle de vie avant de le quitter pour mener une vie non parasitaire, et les parasites permanents qui vivent tout au long de leur existence sur ou dans l’hôte à l’exception du moment où ils mènent une vie libre pour changer d’hôte. Les parasites qui sont égarés sur les hôtes ou endroits inhabituels sont désignés par des termes indiquant la nature de leur anormalité : on définit ainsi les parasites accidentels, qui sont présents occasionnellement sur des hôtes inhabituels dus à des conditions naturelles, et lesparasites aberrants ou erratiques qui sont localisés sur des parties inhabituelles chez unhôtenormal. Toutes les catégories de parasites décrites précédemment dans ceparagraphe sont des parasites obligatoires, incapables de vivre sans effectuer une partie de leur développement sur ou dans l’hôte. Cependant, certains parasites à vie normalement libre peuvent exister pour une courte période dans le corps d’un autre animal, ou accidentellement introduits, ou profiter occasionnellement d’un autreorganisme, comme c’est le cas des mouches Milichiidae qui accaparent les proies déjàcapturées par les invertébrés prédateurs. Ces parasites sont appelés parasites facultatifs.
Enfin, le nombre d’hôtes utilisés par les parasites durant leur cycle de vie permet de les subdiviser en deux principaux groupes, les parasites monoxènes et les parasites hétéroxènes. Les parasites monoxènes n’ont qu’un seul hôte durant toute leur vie, il s’agit d’un hôte définitif. Par opposition, les hétéroxènes utilisent plusieurs hôtes successifs : l’hôte où se déroule le stade adulte est l’hôte définitif et les autres sont des hôtes intermédiaires. Les parasites hétéroxènes qui fréquentent deux hôtes sont dits dixènes,ceux qui fréquentent trois hôtes sont des trixènes, etc.
Ectoparasites des chauves-souris
Les ectoparasites des chauves-souris comprennent une grande diversité d’arthropodes, constitués essentiellement d’arachnides et d’insectes (Whitaker, 1988).
Les arachnides inféodés aux chiroptères comprennent un nombre diversifié d’acariens (Ordres des Astigmata, Mesostigmata, Oribatida et Prostigmata) y compris les tiques (Ordre des Ixodida). Les insectes parasites de chauves-souris comprennent deux familles de mouches (Diptera: Nycteribiidae et Streblidae), une famille de puces (Siphonaptera: Ischnopsyllidae) et une famille de punaise (Heteroptera: Cimicidae).
Les ectoparasites de chauves-souris se caractérisent par une remarquable diversité au niveau de leur cycle de vie, leur écologie, et les adaptations leur permettant de maintenir des relations durables avec leurs hôtes. Alors que certaines espèces appartenant à certaines familles sont des parasites obligatoires, passant leur cycle de vie entier sur le corps des chauves-souris (cas des tiques), d’autres vivent dans le gîte et peuvent infester périodiquement leurs hôtes (cas des Cimicidae). En outre, certaines familles sont considérées comme hautement spécifiques à l’hôte, généralement parasitant une seule espèce de chauve-souris (par exemples les tiques, les Nycteribiidae et les Streblidae), tandis que d’autres familles sont généralistes et moins fidèles à une espèce hôte, pouvant même parasiter d’autres mammifères (cas de certaines espèces de Cimicidae). Les ectoparasites de chauves-souris dépendent également du milieu où vit la chauve-souris qu’ils utilisent comme hôte. En effet, certaines familles sont principalement associées à des espèces de chauves-souris qui gîtent dans les arbres ou des bâtiments (par exemple les puces), d’autres préférant les hôtes cavernicoles (par exemple les Nycteribiidae). Il s’agit en quelque sorte d’une double spécificité parasitaire en termes d’habitat (Milieu) et de biotope (Hôte).
Mouches ectoparasites de chauves-souris
Les Nycteribiidae et les Streblidae, communément appelées « bat flies » (anglais), sont des mouches (Insecta: Diptera) ectoparasites obligatoires, fortement spécialisées et associées uniquement aux chauves-souris (Dick & Patterson, 2006). Elles vivent dans la fourrure et sur le patagium des ailes des chauves-souris où elles consomment du sang. Comme la plupart des parasites obligatoires, la vie des mouches ectoparasites de chauves-souris est étroitement liée à celle de leur hôte, par conséquent leur répartition géographique reflète celle de leurs hôtes (Wenzel et al., 1966; Wenzel, 1976). Généralement, la diversité spécifique de ces mouches est élevée dans les pays tropicaux, moins élevée dans les régions sub-tropicales et plutôt faibles dans les régions tempérées. Plus de 500 espèces de mouches parasitant les différents genres de chauves-souris ont été décrites dans le monde (Dick & Graciolli, 2008; Graciolli & Dick, 2008).
La biologie de la reproduction des Nycteribiidae et des Streblidae demeure actuellement mal connue mais ces mouches se reproduisent majoritairement par viviparité et sont pupipares (Hagan, 1951) : l’œuf est fécondé intérieurement et tous les stades larvaires se déroulent dans la chambre génitale de la femelle adulte, la larve est nourrie dans cette chambre par la sécrétion de glandes spéciales. Ainsi, une femelle gravide porte une seule larve à l’intérieur de son abdomen qui y mue deux fois pour atteindre le troisième stade larvaire. La larve, appelée également prépupe, est ensuite déposée sur le substrat du gîte des chauves-souris, puis la pupaison a lieu, pendant laquelle la peau de la prépupe se durcit pour constituer le puparium et forme ainsi la pupe. Le stade pupal dure environ trois à quatre semaines (Ching & Marshall, 1968) et une mouche adulte émerge et peut coloniser un hôte. La déposition de la prépupe directement sur l’hôte peut avoir lieu, mais les pupes sont déformées et généralement le développement n’arrive pas à son terme. De plus, la plupart des pupes sont nettoyées par l’hôte. Les mouches adultes ne quittent généralement leur hôte que lorsque la femelle dépose la larve sur la paroi du gîte diurne utilisé par les chauves-souris infestées.
METHODOLOGIE GENERALE
Etudes sur le terrain
Site et périodes d’études
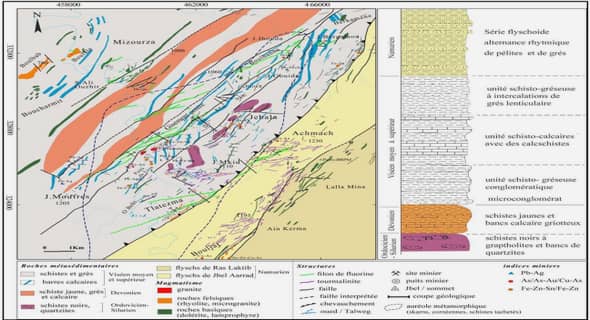
Les échantillonnages de Rousettus madagascariensis ont été effectués dans la Grotte des Chauves-souris (12°57,4’ de latitude Sud et 49°07,1’ de longitude Est, altitude 100 m) dans la partie Est du Parc National d’Ankarana, à 3 km sur route au nord-ouest du village de Mahamasina (Figure 5). Cette grotte héberge une colonie importante de R. madagascariensis (Cardiff et al., 2009; Noroalintseheno Lalarivoniaina, 2015) (Figure 6).
Cette espèce a occupé en permanence la grotte depuis plusieurs décennies ; aucun détail précis n’est connu sur les migrations locales ou régionales vers d’autres gîtes, mais celles-ci ont sûrement lieu. Dans la grotte, la colonie de Rousettus est distribuée en plusieurs groupes sur la voûte de la grotte, cette dernière étant caractérisée par des formations calcaires légèrement humides
Table des matières
INTRODUCTION GENERALE
Chapitre I. GENERALITES
I. 1. Généralités sur les chauves-souris
I. 1. 1. Bio-écologie et taxonomie
I. 1. 2. Chauves-souris de Madagascar
I. 2. Généralités sur les parasites
I. 2. 1. Nature du parasitisme
I. 2. 2. Différents groupes de parasites
I. 2. 3. Ectoparasites des chauves-souris
I. 3. Mouches ectoparasites de chauves-souris
I. 3. 1. Nycteribiidae
I. 3. 2. Streblidae
I. 4. Présentation de l’espèce hôte : Rousettus madagascariensis
I. 5. Caractéristiques du milieu d’étude : Le Parc National d’Ankarana
I. 5. 1. Localisation géographique
I. 5. 2. Géologie et géomorphologie
I. 5. 3. Climat
I. 5. 4. Végétation
I. 5. 5. Chauves-souris d’Ankarana
Chapitre II. METHODOLOGIE GENERALE
II. 1. Etudes sur le terrain
II. 1. 1. Site et périodes d’études
II. 1. 2. Méthodes de capture
II. 1. 3. Identité des individus capturés et méthode de marquage
II. 1. 4. Méthode de détermination de l’âge
II. 1. 5. Méthode de détermination du sexe
II. 1. 6. Méthode de mensuration
II. 1. 7. Méthode de pesage
II. 1. 8. Méthode de collecte des mouches ectoparasites
II. 2. Etude au laboratoire
Chapitre III. DIVERSITE, STRUCTURES DE LA COMMUNAUTE ET DE LA POPULATION DES MOUCHES ECTOPARASITES (DIPTERA: NYCTERIBIIDAE, STREBLIDAE) DE Rousettus madagascariensis (CHIROPTERA: PTEROPODIDAE)
III. 1. Introduction
III. 2. Méthodologie
III. 2. 1. Collecte et nature des données
III. 2. 2. Analyses des données
III. 3. Résultats
III. 3. 1. Résultats de captures
III. 3. 2. Diversité des mouches ectoparasites
III. 3. 3. Structure de la communauté de mouches ectoparasites
III. 3. 4. Structure de la population des mouches ectoparasites
III. 4. Discussion
III. 4. 1. Diversité
III. 4. 2. Structure de la communauté de mouches ectoparasites
III. 4. 3. Structure de la population des mouches ectoparasites
III. 5. Conclusion
Chapitre IV. PARASITISME DES MOUCHES ECTOPARASITES (DIPTERA), Eucampsipoda madagascarensis (NYCTERIBIIDAE) ET Megastrebla wenzeli (STREBLIDAE), CHEZ Rousettus madagascariensis (CHIROPTERA: PTEROPODIDAE) : MISE EN EVIDENCE DES FACTEURS INTRINSEQUES DE L’HOT E ET DE LASAISONNALITE A ANKARANA
IV. 1. Introduction
IV. 2. Méthodologie
IV. 2. 1. Collecte et nature des données
IV. 2. 2. Indices de parasitisme
IV. 2. 3. Analyses des données
IV. 3. Résultats
IV. 3. 1. Prévalence et taux de parasitisme au sein de la population hôte
IV. 3. 2. Influences des facteurs intrinsèques de l’hôte et de la saisonnalité sur l’abondance parasitaire
IV. 4. Discussion
IV. 4. 1. Prévalence et taux de parasitisme au sein de la population hôte
IV. 4. 2. Influence de l’âge et du sexe de l’hôte
IV. 4. 3. Importance de la surface corporelle de l’hôte
IV. 4. 4. Influence de l’activité des glandes cervicales de l’hôte
IV. 5. Conclusion
Chapitre V. DISCUSSION GENERALE
CONCLUSION GENERALE
REFERENCES BIBLIOGRAPHIQUES
GLOSSAIRE
ANNEXES
![]()