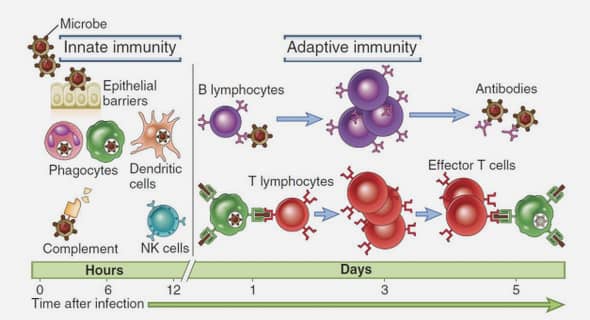
Epidémiologie moléculaire des encéphalites virales aiguës
Les herpèsvirus
Membres de la famille des Herpesviridae, les herpèsvirus sont des virus à ADN (acide désoxyribonucléique) enveloppés, avec une capside icosaédrique. Le mot «herpès» vient du grec «herpein» qui signifie ramper comme un serpent. Les huit herpèsvirus strictement humains sont répartis en 3 sous-familles : les Alphaherpesvirinae (les herpèsvirus simplex de types 1 et 2 (HSV-1 et HSV-2) et le virus de la varicelle et du zona (VZV)), les Betaherpesvirinae (cytomégalovirus (CMV) et les 6e et 7e herpèsvirus humain (HHV-6 et HHV-7)) et les Gamaherpesvirinae (virus Epstein-Barr (EBV) et le 8e herpèsvirus humain (HHV-8)). Ainsi, après une primo-infection ces virus herpétiques se maintiennent dans l’organisme par « une infection latente » et se mettent à l’abri du système immunitaire et des antiviraux. Ils restent ainsi dans l’organisme pour pallier à leur fragilité dans le milieu extérieur. La réactivation de l’infection latente peut conduire à une réinfection endogène, pouvant aller d’une forme souvent asymptomatique à des complications neurologiques ou systémiques graves (Figure 1). Figure 1: Infection latente des herpèsvirus humains dans l’organisme (Source: cours de virologie Université Pierre et Marie Curie, 2006-2007)
Les virus de l’herpès simplex
L’herpès simplex a été identifié dans plusieurs études comme la plus fréquente cause d’hospitalisation chez des patients atteints d’encéphalite, avec une incidence annuelle entre 1/250.000 et 1/500.000 (Honnorat et al., 2013; Vora et al., 2014). L’encéphalite à herpès simplex est due à une infection du SNC par les virus de l’herpès simplex HSV-1 ou le HSV-2. C’est une affection rare pouvant être due à une infection primaire ou récurrente par un HSV, possible chez des patients immunodéprimés. La transmission interhumaine est directe. Le HSV-1 est de loin la plus fréquente cause d’encéphalite infectieuse dans le monde. Il est majoritairement retrouvé chez l’adulte et l’adolescent, constituant près de 90% des cas d’encéphalite à HSV (Whithley, 2006). Après une primo-infection, le HSV-1 reste latent dans le bulbe olfactif et le ganglion de Gasser (Zhang et al., 2007). Une récurrence d’herpès labial est caractéristique de la réactivation endogène de l’infection dans les vésicules mais le virus très neurotrope n’est pas retrouvé dans la peau et les muqueuses. Toutefois, le HSV-2 est associé de façon récurrente à des lésions génitales chez l’adulte et se transmet par voie sexuelle ou congénitale. Chez le nouveau-né, il peut causer des complications allant d’une maladie systémique grave à des encéphalites acquises par voie congénitale avec un taux de mortalité d’environ 50% (Corey & Wald, 2009). Les HSV sont les plus communes causes d’encéphalite identifiées à travers le monde. Entre 2004 et 2014, près de 69% de cas d’encéphalite à HSV ont été décrits en Turquie (Sili et al., 2014). En 2015, six patients atteints d’une encéphalite à HSV-2 ont été également décrits en Gréce (Zis et al., 2016). Des cas isolés ont été également reportés en 2013 en Angleterre (Dodd et al., 2015) et en 2015 dans d’autres pays comme l’Inde, la France, et la Suisse (Kaeley et al., 2016; FernandezGerlinger et al., 2015; Jaques et al., 2016). En Algérie, un taux d’environ 2% de cas d’infection à HSV a été décrit au cours de l’année 2015 (rapport IPA, 2015). Au Sénégal, il n’existe pas de surveillance active des infections à HSV. Cependant, dans le cadre d’une investigation menée à l’institut Pasteur de Dakar (IPD) sur des sérums collectés en 2014, des traces d’une infection à HSV avaient été identifiées chez 1% des échantillons testés (CNRrougeole IPD, 2016)
Le virus de la varicelle ou zona
Le virus de la varicelle et du zona (VZV) est responsable d’une inflammation aiguë et douloureuse des ganglions nerveux avec des éruptions cutanées formant une ceinture autour du corps. Le VZV peut se transmettre par aérosol, par voie directe ou lors de l’accouchement (Nichols et al., 2011; Ahn et al., 2016). Cette maladie dure généralement entre 7 et 10 jours et s’observe le plus souvent chez les enfants. Cependant, le VZV latent peut se réactiver et causer des éruptions vésiculaires, en particulier chez les personnes âgées et les individus immunodéprimés. En effet, comme les autres herpèsvirus, après une primo-infection, le VZV reste latent dans les neurones des ganglions sensoriels. Il peut être responsable d’un large spectre de complications neurologiques dont une névralgie post-herpétique, une myélite, une vasculopathie focale ou une encéphalite. L’exposition aux rayons solaires ultraviolets (UV) pourrait faciliter la réactivation (Murata et al., 2010; Korostil & Regan., 2016). L’incidence de l’encéphalite à VZV varie de 1,3 à 4,8 cas par 100.000 personnes par an (Hope-Simon, 1965) et peut parfois atteindre des proportions plus importantes que les HSV. Les taux d’hospitalisation varient entre 2 et 25 cas par 100.000 personnes par an, avec des proportions plus importantes chez les personnes âgées. Cependant, cette incidence a considérablement diminuée dans certains pays après l’introduction de la vaccination contre la varicelle chez les enfants de moins de vingt-quatre mois (Kawai et al., 2014; Gabutti et al., 2016). Peu de données existent sur l’épidémiologie du VZV en Afrique (Hussey et al., 2016). Au Sénégal, une épidémie de VZV a été enregistrée en juin 2016 avec un taux d’infection de 51,1% (BEH-MSAS, CNR-rougeole IPD, 2016).
Le cytomégalovirus
Le cytomégalovirus (CMV) est l’un des principaux agents viraux responsables d’infections congénitales dans les pays développés, conduisant à des troubles neurologiques chez l’enfant. En effet, durant le premier trimestre de la grossesse, le CMV a un potentiel tératogène et peut causer des malformations du cerveau chez le nouveau-né. Sa prévalence globale varie entre 24 0,2% et 2,5% chez tous les nouveau-nés (González-Sánchez et al., 2015; Xue et al., 2015). Cependant, chez les adultes, la majorité des encéphalites à CMV a été observée chez des personnes immunodéprimées, comme celles atteintes du VIH (virus de l’immunodéficience humaine) (Katchanov et al., 2014). En effet, l’infection à CMV a une prévalence élevée partout dans le monde et les taux d’infection augmentent avec l’âge et la séropositivité qui est estimée entre 40 à 100% chez les individus âgés de plus de cinq ans (Lopo et al., 2011). Les infections à CMV sont fréquemment associées à un risque élevé de mortalité et de morbidité, surtout après une transplantation d’organes (Simon et al., 2016). La primo-infection est en générale asymptomatique ou cause un syndrome grippal. Les épisodes de réactivation sont rapidement contrôlées par la réponse immunitaire de l’hôte (Parry et al., 2016). Néanmoins, une encéphalite est observée dans moins de 1% des infections à CMV (Granerod et Crowcroft., 2007). En Afrique, le CMV a une forte prévalence (Bates & Brantsaeter., 2016). Au Sénégal, il n’existe aucune surveillance des infections à CMV, mais un faible taux d’infection de 1,37% a été enregistré en 2015 dans le cadre d’une investigation effectuée sur des sérums prélevés en 2014 par l’Institut Pasteur de Dakar (CNR-rougeole IPD, 2015).
Les 6e et 7e herpèsvirus humains
Le 6e herpèsvirus humain (HHV-6) est un agent viral ubiquitaire impliqué dans un nombre élevé de cas d’encéphalites virales, y compris des encéphalites aiguës. La primo-infection à HHV-6 survient en général lors de la petite enfance et se manifeste par une maladie fébrile ou des éruptions cutanées bénignes, avec une séroprévalence supérieure à 95% après l’âge de deux ans. Bien que la primo-infection soit asymptomatique, il peut causer de rares complications dont l’exanthème. Comme les autres herpèsvirus, le HHV-6 peut rester latent après une primo-infection dans le cerveau, les reins, les glandes salivaires ou les cellules telles que les lymphocytes T, les cellules germinales de la moelle osseuse ou les cellules de la microglie (Bhanushali et al., 2013). Cependant, la réactivation du HHV-6 chez des patients immunodéprimés, comme ceux atteints du VIH ou ayant subis une transplantation d’organes ou de cellules souches hématopoïétiques, peut conduire à des manifestations allant d’une forme systémique sévère, caractérisée par des crises et des changements de personnalité, à des 25 complications neurologiques parfois fatales (Ogata, 2016). L’encéphalite à HHV-6 peut atteindre de fortes incidences, comprises entre 11,6% et 21,4%, chez les patients ayant subis une transplantation (Ogata, 2015). Ainsi, les transplantations de cellules de la moelle osseuse, de cellules souches du sang périphérique ou de cellules sanguines du cordon ombilical représentent des facteurs de risque. En outre, le HHV-6 est le seul herpèsvirus humain capable d’une intégration chromosomique dans les télomères. Cette intégration chromosomique de l’ADN viral s’observe dans environ 2% de la population humaine (Kaufer & Flamand., 2014; Bhanushali et al., 2013). Si cette intégration se passe dans les cellules de lignée germinale, le HHV-6 intégré de façon chromosomique peut affecter la progéniture par transmission directe et se retrouver ainsi dans toutes les cellules nucléées de leur organisme (Hill et al., 2016). De même, le 7e herpèsvirus humain (HHV-7) présente un profil d’expression et un tropisme similaires à ceux du HHV-6. En effet, le HHV-7 peut réactiver le HHV-6 latent et peut participer au tableau clinique sévère associé à l’infection à HHV-6. Toutefois, le potentiel pathogénique du HHV-7 reste incertain (Staheli et al., 2016). Contrairement au CMV, les autres Béta-herpèsvirus présentent une faible prévalence en Afrique sub-saharienne (Tembo et al., 2015), y compris au Sénégal où un taux d’infection inférieur à 1% a été observé lors d’une étude menée par l’Institut Pasteur de Dakar en 2014 (CNR-rougeole IPD, 2015).
Le virus Epstein-Barr
Le virus Epstein-Barr (EBV) infecte plus de 95% de la population mondiale et cause des mononucléoses infectieuses bénignes avec des symptômes classiques comme une fatigue, une pharyngite et une lymphadénopathie. L’EBVa été le premier virus identifié dans une tumeur humaine; c’est-à-dire le lymphome de Burkitt (Houldcroft & Kellam., 2015). Il a également été associé au lymphome de Hodgkin, au carcinome nasopharyngé et à quelques cancers gastriques (Peng et al., 2016). En effet, l’infection à EBV est associée à des lymphomes du SNC pouvant être accompagnées de complications neurologiques (Weinberg et al., 2002; Hussain RS. & Hussain NA., 2013). Ces formes graves ne s’observent que chez 1 à 18% des patients atteints d’une mononucléose infectieuse et peuvent s’accompagner de manifestations telles que des crises, une ataxie, une encéphalite, un syndrome de Guillain-Barré et une 26 dépression (Jenson, 2011). Comme pour le HSV-1, la transmission de l’EBV est directe à travers la salive et il affecte majoritairement les jeunes adultes. Cependant, l’allaitement est une voie potentielle de transmission du EBV chez les enfants (Daud et al., 2015). Après une primo-infection, l’EBV reste en état de latence dans les cellules épithéliales. Bien que les complications neurologiques apparaissent le plus souvent durant l’infection aiguë, la réactivation de l’EBV latent peut être grave chez des patients immunodéprimés (Cohen, 2000). L’EBVest responsable de 2% à 5% des cas d’encéphalites et de méningites virales et les patients souffrant d’une encéphalite à EBV peuvent présenter un tableau clinique allant d’une fièvre à un stade de coma (Akkoc et al., 2016). L’infection à EBV peut entraîner une large gamme de maladies chez les patients recevant une greffe allo-génique de cellules souches hématopoïétiques, avec une incidence pouvant varier entre 3,3% et 15,6% (Xuan et al., 2013). La séropositivité du donneur en anticorps IgG anti-EBV représente un facteur de risque pour une réactivation du EBV latent qui est associée en général à une encéphalite (Jaskula et al., 2010). Au Sénégal, une étude réalisée en 2014 à l’Institut Pasteur de Dakar a montré un taux de 5,76% d’infections à EBV (CNR-rougeole IPD, 2015)
Le 8e herpèsvirus humain
Comme l’EBV, le 8e herpèsvirus humain (HHV-8) est un agent ubiquitaire responsable de maladies multifactorielles comme le lymphome primaire des séreuses, la maladie multicentrique de Castleman et le sarcome de Kaposi (SK). En effet, il est responsable de toutes les formes du SK qui est le cancer le plus fréquemment associé à l’infection au VIH en Afrique (Courville et al., 2014). Le sarcome de Kaposi affecte plus les hommes que les femmes et existe sous quatre formes : une forme classique retrouvée en Europe de l’Est et sur les contours de la méditerranée, affectant des hommes âgés de 50 à 70 ans ; une forme endémique touchant des sujets plus jeunes en Afrique subsaharienne ; une forme iatrogénique associée aux patients recevant un traitement immunosuppresseur, suite à une transplantation d’organes ; et une forme épidémique liée au VIH (Begré et al., 2016 ; Njiki et al., 2015). Comme l’EBV, le HHV-8 a un potentiel oncogène et est associé à des troubles lymphoprolifératifs, surtout chez les jeunes adultes infectés par le VIH chez qui il peut entraîner des infections neurologiques (Jha et al., 2015). Le HHV-8 possède une distribution 27 mondiale, avec une forte prévalence en Afrique et au Moyen-Orient, une prévalence moyenne en Méditerranée et une faible prévalence aux Etats-Unis et au Nord de l’Europe. Cependant, son mécanisme d’action et son mode de transmission restent inconnus. L’importance du type de transmission varie également selon la région. Aux Etats-Unis et au Nord de l’Europe, la transmission sexuelle entre les homosexuels est la principale voie de transmission. Par contre, dans les zones à forte prévalence, la transmission à travers la salive durant l’enfance est dominante. En outre, l’homosexualité masculine, l’absence de partenaire stable, les rapports bisexuels non protégés chez les prostituées et la toxicomanie représentent des facteurs de risque (Zhang et al., 2014). Peu de données existent sur le HHV-8 au Sénégal où le sarcome de Kaposi est rare, même dans la population de sujets porteurs du VIH. Cependant, une étude épidémiologique réalisé en 2001 a décrit une séroprévalence de 14,3% pour le HHV-8 chez les femmes enceintes (Gaye-Diallo et al., 2001). Cette séroprévalence est relativement faible par rapport aux résultats de pays africains tels que le Cameroun (51%), l’Ouganda (30 à 50%) et la Zambie (60%) qui ont en commun la particularité de se trouver en zone d’endémie du sarcome de Kaposi (BESTETTI et al., 1998). Mais elle est aussi de loin inférieure à la prévalence observée en Gambie où le HHV-8 affecte environ 79% des femmes enceintes. (ARYOSHI et al., 1998).
LISTE DES ABRÉVIATIONS |