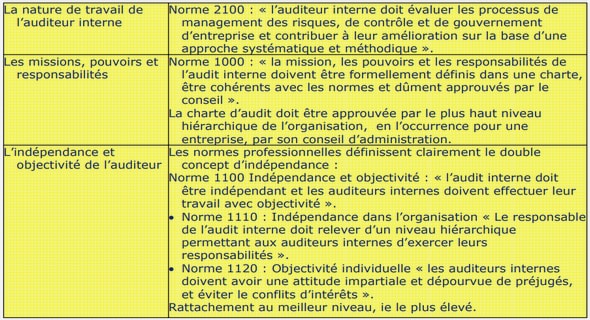
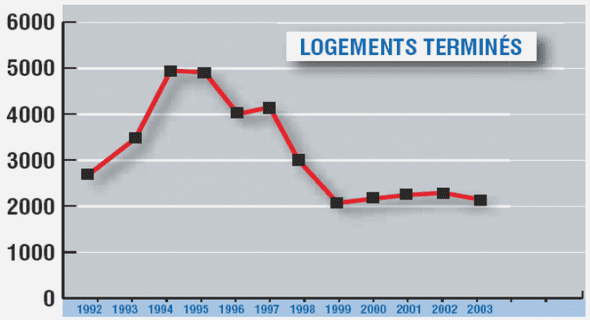
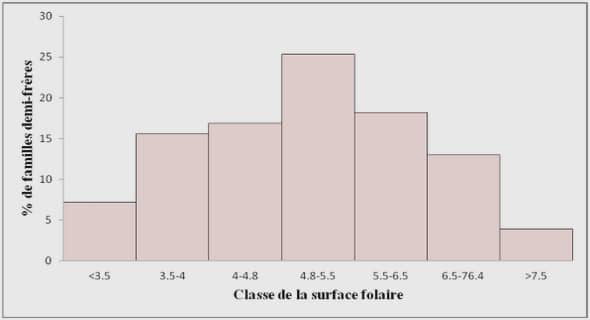
Télécharger le fichier original (Mémoire de fin d’études)
Structure membranaire dans la cellule
Composition
Un des modèles de membrane biologique le plus couramment admis, le modèle de mosaïque fluide (fig. 6), a été décrit par SJ Singer [Singer, 1972]. Dans ce modèle, les lipides constituent une bicouche plane, qui sert de matrice aux protéines membranaires. Lipides et protéines sont libres de mouvements latéraux dans le plan de la bicouche (d’où le terme « fluide »), ainsi que de mouvements de rotation axiale sur eux-mêmes. Le coefficient de diffusion latérale moyen des lipides dans une large variété de membranes biologiques est de l’ordre de [Berg et al., 2002] (notons que cette valeur est de l’ordre de 10 fois plus grande pour des membranes modèles [Lahda et al., 1996]), un lipide peut ainsi visiter la surface de la membrane plasmique d’une cellule en quelques minutes. Pour les protéines, le coefficient de diffusion latérale peut varier sur plusieurs ordres de grandeurs selon sa taille et son affinité et son ancrage dans les lipides de la membrane. Par exemple, il est de l’ordre de pour une rhodopsine [Wey et al, 1981] et de l’ordre de pour une fibronectine ancrée à la membrane plasmique par des filaments d’actine [Berg et al., 2002]. Notons que le coefficient de diffusion latérale dépend de la composition de la membrane. Les mesures diffèrent d’ailleurs selon qu’elles sont effectuées sur membranes modèles ou sur cellules. Une plus forte concentration de protéines a notamment tendance à faire décroitre ce coefficient [Jabobson et al., 1987].
En revanche, les mouvements de lipides ou de protéines d’un feuillet vers l’autre (appelés flip-flop ou diffusion transverse) sont restreints (pour un phospholipide, on compte en moyenne un flip-flop en plusieurs heures [Smith et Green, 1974]), ce qui induit la possibilité d’asymétrie de composition entre les deux feuillets de la membrane. Ce modèle, qui a maintenant plus de quarante ans reste pourtant pertinent dans la plupart des études menées sur les membranes [Nicolson, 2013].
Figure 6 : Modèle membranaire de la mosaïque fluide. Lipides et protéines sont libres de déplacements latéraux dans le plan de la membrane tandis que les mouvements d’un feuillet à l’autre sont fortement restreints. [Nelson et Cox, 2008]
Les lipides constituant les membranes biologiques sont extrêmement variés : il existe plusieurs natures de tête polaire, des chaines carbonées de longueurs différentes et plus ou moins insaturées. Une classification des lipides en 7 catégories est généralement admise [Fahy et al., 1994].
o Les mono-, di- ou triglycérides (aussi nommés acylglycérols) selon le nombre d’acides gras qui estérifient le glycérol, sont les lipides constituant les graisses animales et végétales et permettent de stocker l’énergie métabolique. Leur dégradation permet la production de grandes quantités d’ATP. En moyenne, les glycérides permettent de stocker 38 kJ d’énergie par gramme de lipides [Berg et al., 2002].
o Les glycolipides sont des monosaccharides estérifiés ou amidifiés par des acides gras. Ils se situent généralement sur la face externe des membranes cellulaires chez les eucaryotes et ont pour rôles principaux de faciliter les interactions entre cellules et de stabiliser les membranes lipidiques [Voet et al, 2013].
o Les polycétides sont chimiquement très variés : ils comprennent des macrolides (macrocycles de lactone liés à des sucres) mais également des polyènes, des polyéthers ou des tétracyclines. Il s’agit de métabolites secondaires, c’est-à-dire de composés non directement impliqués dans le développement ou la reproduction d’un organisme, mais qui lui sont néanmoins indispensables à long-terme. Il peut par exemple s’agir de toxines produites par un végétal [Huffman et al., 2010], d’un antibiotique [Robinson, 1991] ou même d’un insecticide.
o Les lipides prénoliques sont formés à partir de l’isoprène. Plusieurs sous-classes dérivées font partie des prénols, comme les caroténoïdes qui agissent comme anti-oxydants ou comme précurseurs de la vitamine A [Rao et Rao, 2007], ou comme les quinones dont dérivent notamment les vitamines E et K [Brunmark et Cadenas, 1989].
o Les sphingolipides sont des lipides dont le squelette est celui d’une sphingosine (un acide gras lié à 2 fonctions alcool et une fonction amine) et qui est lié à d’autres acides gras, et éventuellement, à d’autres groupements comme des sucres ou des groupements phosphate. Ils sont présents, entre autres, dans les membranes plasmiques. Ils ont un rôle important dans la transmission du signal impliquée dans la croissance des cellules, dans la différentiation de ces dernières, dans l’apoptose ainsi que dans la reconnaissance des cellules par le système immunitaire [Bartke et Hannun, 2009].
o Les stérols sont composés d’un noyau stérane avec un groupe OH sur le carbone
o Les phospholipides sont constitués d’un groupe acide phosphorique lié à une molécule polaire (la partie hydrophile) ainsi qu’à, généralement, deux chaines aliphatiques (la partie hydrophobe). Le nombre d’insaturations le long de ces chaines carbonées modifie la mobilité des lipides dans la membrane : une insaturation forme un coude plus rigide qu’une liaison simple et nuit donc à l’alignement des lipides, augmentant ainsi la fluidité générale de la membrane [Kates et al., 1984]. Les phospholipides, avec les sphingolipides décrits précédemment, sont les principaux constituants des membranes biologiques. Ils participent également au métabolisme et à la signalisation cellulaire. Puisqu’il en sera question dans les études décrites dans ce manuscrit et qu’ils sont parmi les plus présents dans les membranes de cellules eucaryotes, citons parmi eux les phosphatidylcholines (PC), les phosphatidylethanolamines (PE) ou encore le diphosphatidyl-glycérol (ou cardiolipide).
Les protéines sont les autres constituants des membranes biologiques. Il s’agit de macromolécules, formées de chaines d’acides aminés liés entre eux par des liaisons peptidiques. Selon les organites, le ratio entre protéines et lipides est différent : de l’ordre de 50% (en masse) de protéines dans la membrane plasmique [Cooper et Hausman, 2004], entre 15 et 30% de protéines dans les membranes myélinisées isolant les axones des cellules nerveuses [Morell et Quarles, 1999]. Dans le cas de la membrane interne des mitochondries, la proportion de protéines est de l’ordre de 70% [Hallermayer et Neupert, 1973 ; Schenkel et Bakovic, 2014]. Les protéines étant des molécules bien plus grosses que les lipides, le rapport protéines sur lipides en nombre de molécules dans une membrane est plutôt de l’ordre de 1 pour 50 [Alberts et al., 1990].
La nature des protéines varie également selon l’organite, avec certaines régions d’un même organite et même entre les deux feuillets d’une même membrane. Les protéines ont des fonctions spécifiques : par exemple, les enzymes catalysent de nombreuses réactions telles que la glycolyse, des canaux protéiques transmembranaires actifs ou passifs transportent ou autorisent la diffusion de molécules spécifiques à travers la membrane, les glycoprotéines interviennent dans la reconnaissance intercellulaire. L’efficacité et le fonctionnement des protéines membranaires dépendent cependant de leur environnement lipidique, de la proximité de certaines autres protéines et de la disponibilité de ressources, facteurs pour lesquels la compartimentation et l’asymétrie de composition membranaire jouent un rôle essentiel.
Compartimentation et asymétrie
L’importance de la compartimentation, c’est-à-dire le fait que plusieurs milieux aux propriétés physiques et chimiques différentes soient définis au sein d’une cellule, ainsi que de l’asymétrie de composition membranaire, s’illustre à travers de nombreuses fonctions des organites cellulaires. Les lysosomes, par exemple, sont de petites vésicules de 0,5 µm de diamètre présents dans les cellules eucaryotes (à l’exception des hématies). Leur membrane permet la séparation de leur milieu interne, dans lequel le pH est maintenu au voisinage de 5 grâce à des pompes à protons membranaires, du cytosol, dans lequel le pH est voisin de 7,2. Les enzymes contenues dans le lysosome, dont le rôle est la digestion intra-cellulaire contrôlée, sont des hydrolases acides, fonctionnant de manière optimale lorsque le pH est de l’ordre de 5. Ainsi, la compartimentation des milieux permet le confinement et l’efficacité des enzymes lysosomales. [Weissmann, 1965 ; Bainton, 1981]
La membrane plasmique a pour rôle premier de délimiter une cellule : la semi-perméabilité de cette membrane assure le maintien, dans le cytosol, du pH et de la concentration de nombreux ions et molécules. La membrane plasmique est enrichie en sphingolipides et en stérols (fig. 7) [Van Meer et al., 2008]. Ces deux lipides sont connus pour augmenter la résistance de la membrane au stress mécanique et pour assurer la préservation de son intégrité [London et Brown, 2000]. Cependant, la composition des deux feuillets de la membrane plasmique est très différente, on parle d’asymétrie de composition : les lipides terminant par une choline (phosphatidylcholine et sphingomyéline) sont préférentiellement situés dans le feuillet externe, quand les lipides dotés d’un groupement aminé terminal (phosphatidyléthanolamine ou phosphatidylsérine) sont préférentiellement situés dans le feuillet interne. Cette asymétrie de composition engendre une asymétrie des charges électriques significative. Cette répartition des charges est nécessaire au fonctionnement de certaines protéines, comme par exemple la protéine kinase C [Newton, 1993], une protéine impliquée dans de nombreuses cascades de signalisation, notamment, dans les mécanismes de la mémoire. Les glycolipides, présents uniquement en surface de la cellule, c’est-à-dire sur le feuillet externe de la membrane plasmique sont un autre marqueur de l’asymétrie de cette membrane [Yamakawa et Nagai, 1978]. Certains de ces glycolipides : les sulfatides et les gangliosides, sont impliqués dans le fonctionnement des pompes à ions sodium-potassium, protéines transmembranaires échangeant les 2 ions en question à contre-gradient à travers la membrane en consommant de l’ATP [Curatolo, 1987]. Les glycolipides sont également des marqueurs de surface [Hakomori, 1975]. Dans les membranes plasmiques d’érythrocytes, les glycolipides sont des sites antigéniques et sont d’une grande importance en médecine car ils interviennent dans la détermination du groupe sanguin du système Kell, système complémentaire aux systèmes ABO et Rhésus [Westhoff et Reid, 2004]. L’asymétrie de composition lipidique de la membrane plasmique prend naissance dans le reticulum endoplasmique, organite des cellules eucaryotes où sont produits la grande majorité des lipides de la membrane plasmique, des membranes mitochondriales, de l’appareil de Golgi… Les phospholipides sont produits dans le feuillet cytosolique du reticulum endoplasmique et des flippases assurent le maintien de la bicouche en translocant des lipides du feuillet interne vers le feuillet externe. Les flippases étant spécifiques d’un type de lipides, elles sont responsables de la distribution asymétrique des lipides dans la membrane du reticulum [Banfield, 2011].
Figure 7 : Composition phopholipidique moyenne des membranes de différents organites [Van Meer et al., 2008]. La composition lipidique est ici exprimée en pourcentage du total des phospholipides chez les mammifères (en bleu foncé) et chez la levure (en bleu clair). Les encarts indiquent le pourcentage molaire de cholestérol et d’ergostérol dans ces membranes.
Compartimentation et asymétrie membranaire sont ainsi essentielles dans la régulation des échanges entre compartiments. Cela est également vrai et particulièrement important pour les membranes de mitochondries.
Les mitochondries
Historique
Des structures, identifiées bien plus tard comme étant les mitochondries, ont été plusieurs fois observées dès 1840, c’est-à-dire quelques années après la découverte du noyau cellulaire. Elles n’ont été reconnues comme formant un même organite qu’en 1890 par Altmann (fig. 8) [Altmann, 1894] qui les appellent alors « bioblastes ».
Il faudra encore attendre 1898 pour que Carl Benda leur donne le nom de mitochondries, du grec « mitos » : fil ou ficelle et « chondros » : granule. Mitochondrie est donc un nom purement descriptif des observations et pour cause, jusqu’en 1934, il n’était possible que d’observer ces « granules en enfilade » au microscope électronique. En 1934 cependant est mis au point un protocole d’extraction d’une fraction de foie de cochon d’inde riche en mitochondrie [Bensley and Hoerr, 1934], ce qui a rendu possible les premières caractérisations biochimiques de cet organite. C’est ainsi que dans les années 1950, le « système de transfert d’électrons » permettant la respiration cellulaire est décrit [Hatefi et al., 1962], de même que la structure des mitochondries.
Description
Figure 9 : (A gauche) Schéma d’une mitochondrie. Illustration : Tutorvista. (A droite) Micrographies électroniques illustrant la diversité et les points communs structurels entre les mitochondries de différents tissus. Barre d’échelle : 200 nm [Vafai et Mootha 2012]
Les mitochondries sont généralement décrites comme des cylindres d’environ 1 µm de diamètre et de 1 à 7 µm de long. Cependant, on sait que ces organites changent de forme constamment : ils se raccourcissent, s’allongent, gonflent, se ramifient et même fusionnent et se séparent d’autres mitochondries [Bereiter-Hahn et Vöth, 1994 ; Hom et Sheu, 2009].
Chaque mitochondrie est délimitée par deux membranes : la membrane mitochondriale externe et la membrane mitochondriale interne (MIM), délimitant ainsi trois milieux : le milieu externe (c’est-à-dire le cytoplasme de la cellule), l’espace intermembranaire (entre les deux membranes), et la matrice mitochondriale (fig. 9).
La membrane externe est constituée de lipides et de protéines dont le rapport en masse est de l’ordre de 0,9 µg de lipides pour 1 µg de protéines [Comte et al., 1976]. La plupart de ces protéines sont des porines, c’est-à-dire des canaux permettant les échanges de petites molécules et ions entre l’extérieur de la mitochondrie et l’espace intermembranaire.
La membrane interne est très différente. Elle est formée de nombreuses convolutions de 30 à 40 nm de large [Lea et al., 1994], appelés crêtes ou cristae, qui micro-compartimentent la membrane interne [Mannella, 2006]. Du fait de ces repliements, la quantité de membrane interne est particulièrement importante : la surface totale de la membrane interne est égale à environ 5 fois la surface de la membrane plasmique [Coopes et Hausman, 2007].
La membrane mitochondriale interne contient 0,5 µg de lipides pour 1 µg de protéines [Comte et al., 1976]. Environ 70% en masse des protéines correspondent aux complexes protéiques de la chaine de transfert d’électrons ainsi qu’à l’ATP-synthétase [Alberts et al., 1990]. Sa composition lipidique diffère également de celles des autres membranes de la cellule. En particulier, elle contient en forte proportion un lipide spécifique : le cardiolipide (de l’ordre de 18%) (fig. 10) [Daum, 1984; Krebs et al., 1979 ; Ren et al., 2014].
Figure 10 : Structure chimique du cardiolipide. En vert sont représentés les atomes de carbone, en rouge les oxygènes, en blanc les hydrogènes, en violet les phosphate et en jaune les contre-ions sodium. Image provenant du site internet de Avanti Polar Lipids.
Le nom « cardiolipide » vient du fait que ce lipide a au départ été découvert dans des cellules cardiaques. On sait aujourd’hui qu’il est présent et produit dans toutes les mitochondries. Le cardiolipide est constitué de deux phosphatidylglycérol liés entre eux par des liaisons ester sur un glycérol. Le cardiolipide comporte ainsi quatre chaines aliphatiques. Ce lipide est responsable de l’imperméabilité très élevée aux ions et aux grosses molécules de la membrane interne mitochondriale, qui reste perméable aux seules molécules de dioxygène, de dioxyde de carbone et d’eau.
Les mitochondries possèdent leur propre ADN, l’ADN mitochondrial, dont le fonctionnement est différent de l’ADN contenu dans le noyau cellulaire. Par exemple, une même séquence de bases code pour des protéines différentes dans le noyau et dans la mitochondrie : UGA est un codon « stop » dans l’ADN du noyau mais code pour du tryptophane dans l’ADN mitochondrial des mammifères [Barell et al., 1979]. D’autre part, les mitochondries se divisent par scissiparité, indépendamment de la division de la cellule [Kuroiwa et al., 1998]. L’ADN mitochondrial et la reproduction par scissiparité sont les arguments forts de l’hypothèse endosymbiotique, selon laquelle les mitochondries proviennent de l’incorporation, au cours de l’évolution, de certaines bactéries ou archées dans des cellules eucaryotes [Margulis, 1975 ; Margulis et Bermudes, 1985]. Un eucaryote, l’amibe Paloxyma Palustris, semble confirmer cette hypothèse car, dépourvue de mitochondrie, elle utilise cependant le métabolisme oxydatif des bactéries avec lesquelles elle est en symbiose [Van Bruggen et al., 1983]. Un autre organisme eucaryote, Monocercomonoides sp., produit son énergie par un mécanisme smiliaire à celui des bactéries et semble montrer une perte des mitochondries au cours de son évolution [Karnkowska et al., 2016].
Production d’ATP et mort cellulaire
Les mitochondries ont un rôle important à la fois dans la vie et dans la mort de la cellule : elles génèrent l’ATP nécessaire au fonctionnement de la plupart des protéines de la cellule, et, sous certaines conditions, initient l’apoptose de la cellule.
Production d’ATP
La production d’énergie sous forme d’ATP dans la cellule est possible par deux voies. La première voie est celle de la glycolyse, c’est-à-dire l’utilisation et la dégradation de molécules de glucose, qui aboutit à la formation de pyruvate (étape 2 de la fig. 11). Mais les produits finaux peuvent encore libérer une grande quantité d’énergie par une oxydation plus poussée : la respiration, qui constitue la seconde voie de production d’ATP. Cette oxydation débute avec le cycle de Krebs et termine par la phosphorylation oxydative (étape 3 de la fig. 11). Le cycle de Krebs est constitué d’une série de réactions chimiques, catalysées par des enzymes présentes dans la matrice mitochondriale, démarrant avec l’acétyl CoA produit notamment à partir du pyruvate issu de la glycolyse.
Ces réactions chimiques produisent, pour la plupart, du , forme réduite du nicotinamide adénine dinucléotide ( ), ou du , forme réduite de la flavine adénine dinucléotide ( ). Le transfert des électrons acquis par ces molécules lors du cycle de Krebs constitue la première étape de la phosphorylation oxydative.
Le principe de cette phosphorylation oxydative est de convertir l’énergie chimique contenue dans des molécules de et de en l’énergie de liaison entre un groupement phosphate et une molécule d’Adénosine Di-Phosphate (ADP). Cette conversion est notamment possible grâce à une chaine de transport d’électrons. Cette chaine est constituée d’une série de complexes protéiques incluse dans la membrane mitochondriale interne. Ces complexes, nommés I, II, III et IV, transfèrent des électrons depuis des espèces donneuses (comme le ) vers des espèces acceptrices. Chaque complexe ayant plus d’affinité pour les électrons que le complexe précédent, les électrons passent en cascade d’un complexe à l’autre, jusqu’à l’accepteur final : l’oxygène (fig. 12). Chacune de ces réactions oxydatives « pompe » des protons de la matrice mitochondriale vers l’espace intermembranaire, en traversant la membrane interne mitochondriale.
Figure 12 : La phosphorylation oxydative. Sont ici présentés les éléments intervenant dans la chaine de transport d’électrons ainsi que l’ATP synthétase en bout de chaine. Les électrons provenant des NADH sont transportés de complexe en complexe jusqu’à l’oxygène. A chaque étape de ce transport, des protons sont pompés de la matrice mitochondriale vers l’espace intermembranaire. Le gradient ainsi établi fournit l’énergie nécessaire au fonctionnement de l’ATP synthétase, qui produit alors l’ATP. [Krauss 2001]
Le gradient électrochimique de protons établi par ce pompage est consommé lors du fonctionnement de l’ATP-synthétase [Mitchell et Moyle, 1967]. Cet important complexe protéique enzymatique (environ 15% en masse des protéines de la membrane interne) assemble une molécule d’ADP (adénosine diphosphate) et un phosphate pour former l’ATP (fig.12). L’ATP-synthétase fonctionne à la manière d’un moteur moléculaire constitué d’un stator (les sous-unités et ) et d’un rotor (les sous-unités et ) dont les 10 sous-unités sont disposées en couronnes (fig. 13). L’entrée d’un proton au niveau du rotor modifie deux des sous-unités qui le compose : l’une est protonée alors que la suivante est déprotonnée. La modification des charges de ces sous-unités entraine une rotation de la couronne du rotor par interaction électrostatique avec le stator [Allison, 1998]. Cette rotation entraine des changements cycliques de conformation des sous-unités constituants le stator et donc des changements cycliques d’affinité de ces protéines envers l’ADP et l’ATP. L’enchainement cyclique des trois conformations possibles des sous-unités du stator catalyse la formation de l’ATP à partir d’ADP et de phosphate en [Nakamoto et al., 1999] :
i) se liant à un ADP et un phosphate
ii) formant l’ATP à partir des deux réactifs
iii) libérant l’ATP produit
Il y a ainsi un couplage entre le transport des protons et la synthèse de l’ATP par l’ATP synthétase.
Figure 13 : A) ATP synthétase. Les sous-unités α et β constituent le stator, les sous-unités c sont transmembranaires et constituent le rotor. Les protons, pompés de la matrice mitochondriale au niveau de la sous-unité a modifient les charges des sous-unités c. Les interactions électrostatiques entre ces dernières et le stator entrainent alors la rotation du rotor. [Nakamoto and al., 1999] B) Cycle de l’ATP synthétase. Des changements successifs de conformation des sous-unités de l’ATP synthétase, couplés à la rotation du rotor, conduisent à la formation d’ATP. Ces changements sont cycliques, le complexe retrouve sa conformation initiale après une rotation complète du rotor. [Lodish et al., 2007]
La phosphorylation oxydative est un processus chimique efficace. Chaque molécule de fournit l’énergie nécessaire pour la production de 3 molécules d’ATP, chaque molécule de pour celle de 2 molécules d’ATP en moyenne. Si on prend en compte toutes les réactions qui produisent de l’énergie à partir d’une molécule de glucose jusqu’à son oxydation complète, ce sont environ 130 molécules d’ATP qui sont créées [Alberts et al., 1990]. Cette quantité d’ATP pourra ensuite être consommée par la cellule, pour le fonctionnement de ses protéines. La réaction de décomposition de l’ATP en ADP et phosphate libère une énergie de l’ordre de par molécule d’ATP [Rosing et Slater, 1972].
Mort cellulaire par apoptose
La mitochondrie est l’un des sites de contrôle du déclenchement de l’apoptose, la mort cellulaire programmée des cellules [Susin et al., 1998]. L’apoptose est une fonction physiologique, une auto-destruction cellulaire contrôlée et nécessaire à l’organisme. Contrairement à la nécrose, durant laquelle la membrane plasmique éclate, l’apoptose conserve l’intégrité de la membrane plasmique de la cellule. A la fin du processus, la cellule est ensuite proprement éliminée par l’organisme : elle expose des lipides (la phosphatidylsérine) constitutifs du feuillet interne de la membrane plasmique et est alors phagocytée par les macrophages évitant ainsi l’inflammation [White, 1999].
La voie mitochondriale de l’apoptose peut avoir pour origine une surexposition à des radiations ou à des toxines, une hyperthermie, une hypoxie, des infections virales ou encore une forte concentration en radicaux libres [Elmore, 2007]. Quand l’intégrité de la membrane mitochondriale ne peut plus être préservée, son oxydation et sa perméabilisation sont les premiers signaux menant à la mort de la cellule (fig. 14). Le gradient de protons établi par le fonctionnement de la chaine respiratoire, du fait de la perméabilité de la membrane, disparait, ce qui participe à faire chuter le potentiel membranaire. Parallèlement, la chaine respiratoire interrompt son fonctionnement. Le cytochrome c, une fois relâché dans le cytosol, s’associe à différentes protéines pour former un complexe nommé apoptosome. La caspase 9, enzyme cytosolique clivant les protéines, est activée par l’apoptosome et peut à son tour activer les autres protéines de la même famille, conduisant ainsi à la dégradation contrôlée des composants cellulaires [Susin et al., 1998 ; Desagher et Martinou, 2000].
Table des matières
1 Introduction
1.1 De l’eau aux bicouches lipidiques
1.1.1 L’eau
1.1.2 Amphiphiles et organisation spontanée
1.1.3 Les lipides
1.2 Structure membranaire dans la cellule
1.2.1 Composition
1.2.2 Compartimentation et asymétrie
1.2.3 Les mitochondries
a) Historique
b) Description
c) Production d’ATP et mort cellulaire
d) Liens entre fonction, structure et cardiolipide
1.3 Oxydation du vivant
1.3.1 Initiateurs de l’oxydation
1.3.2 Stress oxydant
1.3.2.1 Lésions des molécules biologiques
1.3.2.2 Péroxydation des lipides
1.3.2.3 Produits d’oxydation des lipides
1.3.2.4 Mécanismes de régulation cellulaire de l’oxydation
1.4 Objectifs du travail présenté
1.5 Organisation du manuscrit
2 Systèmes étudiés
2.1 Vésicules lipidiques
2.1.1 Membranes biomimétiques
2.1.2 Formation de vésicules lipidiques
2.1.3 Propriétés mécaniques d’une membrane
2.1.4 Tension de membrane et tension de lyse d’une vésicule
2.1.5 Perméabilité d’une membrane
2.1.6 Autres propriétés
2.2 Photo-oxydation de vésicules lipidiques
2.2.1 Photo-sensibilisateur
2.2.2 Chlorine-e6
2.2.3 Production d’oxygène singulet
2.2.4 Le choix du photosensibilisateur
2.2.5 Effets de l’oxydation des lipides sur la morphologie d’une vésicule
2.2.6 Perméabilisation dune vésicule géante par oxydation
Conclusion du chapitre
3 Matériel & méthodes
3.1 Produits
3.1.1 Solutions tampons et solvants
3.1.2 Lipides et membranes
3.1.2.1 Lipides
3.1.2.2 Mitoplastes
3.1.3 Photo-sensibilisateurs
3.1.4 Autres produits
3.2 Préparation des vésicules
3.2.1 Préparation des LUV
3.2.2 Préparation des GUV
3.3 Appareils
3.3.1 Spectroscopie d’absorbance et de fluorescence
3.3.2 Microscopie
3.3.2.1 Matériel
3.3.2.2 Acquisition et traitement des images
3.3.2.3 Analyse
3.3.2.4 Microscopie de fluorescence
3.3.3 Microspectroscopie Raman / Pinces optiques
3.3.3.1 Principe
3.3.3.2 Caractéristiques du montage
3.3.3.3 Traitement des spectres obtenus
3.3.4 Autres appareils
4 Interactions entre la Chlorine-e6 et les membranes lipidiques
4.1 Mesure d’affinité entre la chlorine-e6 et les membranes lipidiques
4.2 Résultats et discussions
4.2.1 Spectres d’émission de fluorescence obtenus
4.2.2 Constantes d’affinité
Conclusion du chapitre
5 Effets du cardiolipide dans la résistance des membranes lipidiques à un stress oxydant
5.1 Conception des membranes
5.2 Transitions de forme et perméabilisation de vésicules subissant un stress oxydant
5.2.1 Effets du cardiolipide sur la dynamique des changements morphologiques
5.2.2 Variations de l’aire membranaire et ouverture de pores durant la phase II
5.2.2.1 Délai entre deux ouvertures de pores
5.2.2.2 Evolution du module élastique et de la tension de membrane lors de la perméabilisation
5.3 Oxydation de membranes composées de PC, PE et cardiolipide
5.3.1 Effets du PE
5.3.2 Comportement d’une IM-GUV oxydée
5.4 Discussions
5.4.1 Origine de l’augmentation de la pression osmotique responsable des cycles de gonflement
5.4.2 Défauts membranaires et perméabilisation
Conclusion du chapitre
6 Influence des insaturations des chaines carbonées dans la résistance des membranes au stress oxydant
6.1 Transitions de forme et perméabilisation
6.1.1 Transitions de forme durant la phase A
6.1.2 Perméabilisation
6.1.3 Module élastique et tension de membrane
6.2 Chimie en temps réel de l’oxydation photo-induite
6.2.1 Pics principaux des spectres Raman des lipides
6.2.2 Evolution temporelle des spectres Raman
6.3 Discussions
6.3.1 Temps de début de perméabilisation
6.3.2 Nécéssité de la délocalisation des électrons
Conclusion du chapitre
Conclusions générales et perspectives
Annexes
A.1 Macro ImageJ utilisée pour le repérage des contours des vésicules
A.2 Macro ImageJ utilisée pour la quantification du contraste