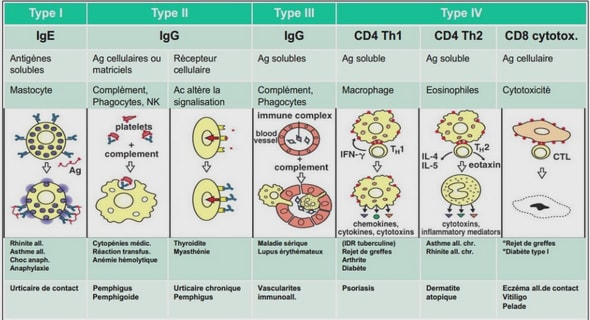
L’ARCHITECTURE GENETIQUE DE LA RESISTANCE A DIFFERENTS PATHOGENES
Détection de QTL de résistance à la nodavirose dans trois populations de bar européen par l’utilisation d’une puce de 57k SNP. Cette section a été publiée en 2020 dans Aquaculture 530 (735930) sous le nom : « Genome-wide association studies for resistance to viral nervous necrosis in three populations of European sea bass (Dicentrarchus labrax) using a novel 57k SNP array DlabChip. » .
Résumé
La Nécrose Nerveuse Virale (NNV) ou nodavirose est une menace majeure pour la production aquacole de bar européen (Dicentrarchus labrax), pour laquelle l’amélioration de la résistance par sélection génétique pourrait être une solution intéressante. Avec le développement des technologies de séquençage haut-débit, l’identification de régions génomiques impliquées dans la résistance est un moyen potentiel d’améliorer l’efficacité de la sélection. L’objectif de cette étude a été d’identifier les régions du génome (Quantitative Trait Loci – QTL) impliquées dans la résistance à la NNV, et de quantifier leurs effets. Quatre familles expérimentales issues de rétro-croisements entre pères hybrides résistants-sensibles et mères sensibles, de 378, 454, 291 et 211 individus ont été produites. Par ailleurs, deux populations commerciales (A et B), de 1 027 et 1 042 individus ont été obtenues à partir de plans de croisement factoriels partiels (59♂ x 20♀ pour la population A; 39♂ x 14♀ pour la population B). Ces six groupes (4 familles et 2 populations commerciales) ont été soumis à un challenge infectieux par balnéation avec un nodavirus de type génétique RGNNV (redspotted grouper nervous necrosis virus). Un panel de SNP (Single Nucleotide Polymorphism) de haute densité a été développé pour créer une puce à SNP (ThermoFisher AxiomTM 57k SNP DlabChip), qui a été utilisée pour le génotypage de tous les individus ainsi que pour la création d’une carte génétique de haute résolution. Dans les quatre familles de rétrocroisement, respectivement 30 917, 23 592, 30 656 et 31 490 marqueurs ont été utilisés dans une approche de cartographie d’intervalles. Dans les populations commerciales, 40 263 marqueurs dans la pop A et 41 166 marqueurs dans la pop B ont permis de réaliser des études d’association pangénomiques (GWAS – Genome-Wide Association Studies) avec deux modèles, GBLUP et BayesCπ. 45 Un QTL a été identifié sur le chromosome LG12 dans trois des quatre familles de rétro-croisement, et un QTL supplémentaire a été détecté sur le LG8, uniquement dans une famille. Chez les populations commerciales, la détection de QTL a révélé un total de sept QTL, parmi lesquels a été retrouvé le QTL positionné sur le LG12, présent dans les deux populations. Ce QTL, qui a été positionné dans un intervalle de confiance de 3,45 cM, explique 9,21% de la variance génétique dans la pop A, alors que les autres QTL identifiés expliquent chacun moins de 1% de la variance génétique. L’identification de QTL impliqués dans la résistance à la nodavirose chez le bar européen, dont un avec un effet fort, devrait avoir un impact important sur l’industrie aquacole. De futurs travaux pourront se concentrer sur la détection précise de la mutation causale du QTL présent sur le LG12 par une approche de séquençage du génome complet.
Introduction
European sea bass (Dicentrarchus labrax) is a major species for Mediterranean aquaculture, with a production of more than 150,000 tons in 2016 (FEAP, 2017). A major threat for the sea bass aquaculture industry is the occurrence of disease outbreaks, and especially of Viral Nervous Necrosis (VNN) disease mostly in warmer growing Mediterranean areas (Breuil et al., 1991; Le Breton et al., 1997; Vendramin et al., 2016). The VNN disease is caused by the Nervous Necrosis Virus (NNV), a RNA virus belonging to the Betanodavirus genus (Mori et al., 1992). NNV is widespread worldwide and can infect more than 70 marine and freshwater species (Doan et al., 2017b). In sea bass production, mortalities up to 90% have been recorded especially at the very susceptible larval and juvenile stages(Le Breton et al., 1997). Several vaccine strategies have been tested, leading to an improvement of survival (Doan et al., 2017b). However, three points limit the use of vaccines in VNN outbreak management: i) vaccines are generally expensive, ii) the number of fish to vaccinate is high which increases the cost issue (Ulmer et al., 2006), and iii) VNN is more infectious in early life stages, while vaccination of small fish is challenging (Gomez-Casado et al., 2011; Sommerset et al., 2005). One promising solution to reduce the effects of VNN outbreaks in aquaculture farms is selective breeding for improved resistance to VNN. Selective breeding for disease resistance in aquaculture species has demonstrated its interest with a genetic gain in survival of 12.5% per generation on average, across different host-pathogen pairs (Gjedrem and Robinson, 2014). For selective breeding to be successful, enough genetic variation for disease resistance within the species is however required. Moderate to high heritability has already been reported for resistance to VNN in European sea bass (from 0.26 to 0.43), confirming the interest of selective breeding for this trait (Doan et al., 2017a; Palaiokostas et al., 2018a). Genomic markers can be used to detect Quantitative Trait Loci (QTL), which are regions of the genome involved in the phenotypic variation of the trait (Georges et al., 1993). Their use allows a better understanding of the genetic architecture of complex traits as well as the location of the potential candidate genes involved in the resistance phenotype. With the access to high-throughput genotyping technologies at a reasonable cost for non-model species, some disease resistance QTLs have been found in a limited number on aquaculture species (see review by Ødegård et al. (2011)). In Atlantic salmon, major QTLs were discovered for IPNV resistance, explaining up to 83% of the genetic variance (Houston et al., 2008; Moen et al., 2009). These results can then be used to accurately choose broodstock among the candidates for selection or commercial diffusion by Marker Assisted Selection (MAS) (Houston et al., 2008; Moen et al., 2009). In Asian sea bass (Lates calcarifer), QTL mapping revealed nine QTLs involved in VNN resistance, explaining between 1.6% and 2.7% of the phenotypic variance (Wang et al., 2017). In Atlantic cod (Gadus morhua), five QTLs were detected, including three 46 QTL with major effects, explaining between 14.2% and 19.7% of the phenotypic variance (Baranski et al., 2010). For VNN resistance in sea bass, a recent study reported three first QTLs, each of them explaining a small part of the genetic variance (1.5% to 4%) (Palaiokostas et al., 2018a). In this study, we investigated the genetic architecture of VNN resistance in sea bass. We performed Genome Wide Association Studies (GWAS) based on two commercial cohorts and four experimental backcross families, all genotyped on the newly developed ThermoFisher AxiomTM Sea Bass 57k SNP DlabChip. Two approaches were considered to detect associations between SNPs and resistance to VNN, interval mapping and a multi-marker Bayesian variable selection model to estimate the sharing of genetic variance explained by the QTLs, as well as their credibility intervals.