Généralités sur les symbioses mycorhiziennes
Les plantes terrestres tirent l’eau et les nutriments du sol. La rhizosphère, partie du sol sous l’influence de la racine et des microorganismes, constitue une niche écologique caractérisée par une forte densité de microorganismes interagissant entre eux et avec les racines.
C’est la zone d’échange entre la plante et le sol environnant. Dans cette rhizosphère, vivent divers microorganismes dont des virus, des bactéries, des actinobactéries, des champignons et des algues (Biais et al, (2006).
Les bactéries et les champignons représentent l’essentiel de la biomasse microbienne du sol (Lavelle et Spain, 2001). Ces microorganismes (bactéries et champignons) interviennent dans les cycles biogéochimiques de la matière (cycles du carbone, du phosphore, etc.) et par la même occasion interagissent directement ou indirectement avec les plantes (symbiotes, phytopathogènes, saprophytes, décomposeurs…). Les bactéries et les champignons sont donc indispensables au maintien de la fertilité et de la productivité des écosystèmes (Coleman et al., 2004).
Certains champignons développent avec les racines des plantes des organes appelés mycorhizes. Du grec «Mikès» (champignon) et du latin «Rhiza» (racine), le terme «mycorhize» fut proposé pour la première fois par le phytopathologiste allemand Frank en 1885. Les mycorhizes désignent des associations symbiotiques contractées par les racines des végétaux avec des champignons du sol. La symbiose mycorhizienne est donc une association mutualiste qui unit un champignon filamenteux aux racines d’une plante supérieure (Smith et Read, 2008).
Le champignon hétérotrophe vis-à-vis du carbone, puise ses ressources énergétiques à partir des photosynthétats de la plante et en contrepartie, le champignon mycorhizien améliore la nutrition hydrominérale de la plante (Smith et Read, 2008). Les champignons mycorhiziens constituent donc une composante essentielle du système sol-plante (Finlay, 2008). Sur la base de critères morphologiques, physiologiques et évolutifs (Smith et Read, 2008), cinq principaux types d’associations mycorhiziennes ont été décrits . Il s’agit des ectomycorhizes, des mycorhizes à arbuscules, des endomycorhizes à pelotons intracellulaires des Orchidaceae, des mycorhizes éricoïdes ou endomycorhizes à pelotons des Ericaceae et des mycorhizes subépidermiques ou ectendomycorhizes.
La symbiose mycorhizienne arbusculaire
Environ 6000 espèces de champignons appartenant au phylum des Glomeromycotina, Ascomycotina et Basidiomycotina ont été enregistrées comme mycorhiziennes (Taktek, 2015). C’est la symbiose terrestre la plus répandue (Smith et Read, 2008), et la plus ancienne dans la nature du fait de leur ubiquité (on les retrouve partout) et du nombre élevé d’espèces végétales concernées (400.000 espèces connues) (Taktek, 2015). La symbiose mycorhizienne arbusculaire pourrait être antérieure au Dévonien et concerne plus de 80 % des plantes vasculaires, soit environ 250.000 espèces de plantes terrestres (Smith et Read, 2008). Il n’existe guère à la surface des continents de formation végétale qui ne comporte pas des plantes à endomycorhizes arbusculaires (Garbaye, 2013). La distribution des espèces de champignons mycorhiziens à arbuscule (CMA) est fortement dépendante de la flore, de l’environnement et de la compétition interspécifique entre plantes (Lekberg et al., 2007). Toutefois, un faible endémisme des CMA a été récemment révélé (Davison et al, 2015). Les CMA regroupés dans le phylum des Glomeromycota, sont des champignons filamenteux coenocitiques qui colonisent à la fois le sol et les racines en établissant un pont entre la rhizosphère et la plante (Dalpé, 2005). Du fait de leur morphologie, leur faible spécificité, et de leur plasticité génétique et fonctionnelle, les CMA sont capables de transporter rapidement des éléments nutritifs entre le sol et la plante hôte ou entre différentes plantes hôtes. En effet, les CMA ne possèdent pas de cloisons, et s’adaptent à des situations variées du fait de la diversité de leur matériel génétique reconfigurable en permanence (Garbaye, 2013). Cette dernière propriété expliquerait en partie comment ce groupe de champignons est resté si stable.
La reproduction des champignons mycorhiziens à arbuscules
Aucune forme de sexualité n’est connue chez les CMA, c’est un groupe de champignons réputés asexués appartenant au phylum des Glomeromycota (Garbaye, 2013). Ils se propagent donc de façon végétative et par reproduction asexuée, principalement par de grosses spores, de l’ordre de grandeur du dixième de millimètre, jusqu’à un demi-millimètre pour certaines espèces. La forme des spores est généralement sphérique, mais chez certaines espèces elles peuvent présenter une ampoule basale ou être accompagnées d’une sorte de sac annexe, dont le contenu initial a contribué à la formation de la spore proprement dite. La classification conventionnelle des CMA repose presque entièrement sur les caractères morphologiques des spores et la structure de leur paroi. Les spores sont produites dans le mycélium externe des CMA, à l’extrémité des hyphes et sont dispersées par les mouvements des particules de terre dus au vent et au ruissellement, mais aussi par le transport actif des petits animaux du sol (Garbaye, 2013).
Les spores de CMA sont capables de germer spontanément, si les conditions physico-chimiques sont favorables, même en absence de plante hôte(Smith et Read, 2008). Cependant, sans la plante hôte, la croissance des hyphes germinatifs sera extrêmement limitée et cessera après quelques jours. Le cytoplasme des hyphes germinatifs se rétractera jusqu‘à retourner dans la spore (Logi et al., 1998). Pendant cette phase le champignon est en vie ralentie, il respire peu et consomme très peu ses réserves. Ce processus permet à la spore de rester en vie et de maintenir sa capacité de germer à nouveau pour éventuellement coloniser les racines d’une plante à proximité (Becard et al., 2004).
Importance des champignons mycorhiziens à arbuscules
Importance dans la nutrition des plantes : L’association symbiotique entre des champignons du sol et les racines de la majorité des plantes terrestres entraine un échange bidirectionnel de composés carbonés de la plante vers le symbionte fongique et de composés minéraux du champignon vers la plante (Dalpé, 2005). Le champignon permet à la plante d’explorer un volume important de sol pour s’approvisionner en eau et éléments minéraux. La symbiose améliore ainsi de manière remarquable la croissance et la nutrition phosphatée des plantes et leur permet d’exploiter au mieux les ressources en eau (Bâ et al., 2001 ;Oehl et al., 2011).
La colonisation mycorhizienne a un impact significatif sur la nutrition des plantes en phosphore, dont la disponibilité dans le sol est un facteur limitant de la croissance des plantes (Helgason et al., 2002). Le P est indispensable pour les cellules vivantes vu qu’il est un composant principal de l’ADN, de l’ARN, de l’ATP et des phospholipides (composant majeur des membranes cellulaires). La flore microbienne du sol joue un rôle majeur dans le cycle du P, car elle établit un lien entre le réservoir de P dans l’environnement vivant et non vivant. Certains microorganismes tel que les CMA facilitent l’altération, la minéralisation et la solubilisation des différentes formes de P pour permettre leur utilisation par les plantes (Taktek, 2015). Pour Duponnois et al., (2012), le rôle majeur des mycorhizes se situe au niveau de la mobilisation pour la plante d’éléments nutritifs peu mobiles dans le sol, principalement le phosphore.
Importance face aux stress abiotiques et biotiques : Les plantes mycorhizées tolèrent mieux les facteurs de stress d’origine biotique et abiotique (Egli et Brunner, 2002 ; Dalpé, 2005). Des études consacrées à l’effet protecteur des mycorhizes vis-à-vis des agents pathogènes telluriques ont montré que certains symbiotes sont efficaces contre la plupart des agents pathogènes du sol, en particulier des parasites fongiques dommageables aux essences forestières (Perrin, 1985). En effet, les mycorhizes possèdent des capacités de protection des plantes contre les agents pathogènes d’origine tellurique. Cette protection s’opère à travers des mécanismes variés, complémentaires, qui agissent généralement de concert. Mais l’existence et l’efficience de ces différents processus varient notablement selon la plante, le champignon mycorhizien, et l’agent pathogène (Perrin, 1985).
Les mycorhizes protègent aussi les végétaux des effets toxiques des polluants. Certains champignons mycorhiziens résistent aux polluants même si leur teneur est élevé dans le sol (Egli et Brunner, 2002). En effet ils jouent le rôle de filtre en accumulant les polluants au niveau de leur fructifications (Egli et Brunner, 2002). Mais il faut noter que les excès du phosphore en particulier, ou d’autres éléments peuvent contribuer à amoindrir la résistance des parties aériennes des plantes à différentes maladies (Perrin, 1985).
Les phosphates naturels (PN)
Les phosphates naturels sont connus pour leurs teneurs élevées en phosphore. Ils sont utilisés soit en tant que matières premières pour la fabrication industrielle d’engrais phosphatés solubles, soit comme source de phosphore pour l’application directe en agriculture. Ces PN contiennent également des minéraux accessoires ou des impuretés et des éléments radioactifs (uranium…), dangereux pour la santé humaine et animale et toxiques à certaines concentrations pour certains végétaux et microorganismes. Toutefois, les teneurs et les combinaisons de ces éléments varient selon les gisements de PN.
Les phosphates naturels sont peu solubles, on les fait donc réagir avec de l’acide sulfurique pour produire de l’acide phosphorique concentré pour la formation d’engrais phosphatés. Les engrais phosphatés minéraux sont dérivés de phosphates naturels, généralement l’apatite. Le Maroc et le Sahara occidentale possèdent 70% des réserves mondiales d’apatite, estimées à 71000 millions de mètre/tonne. Les premiers producteurs de phosphates naturels en 2011 sont la Chine (38%), les Etats Unis (15%), le Maroc (14%) et la Russie (6%) (Ziadi et al., 2013).
Dans le sol, il existe différentes formes de phosphore : le phosphore soluble organique et inorganique, le phosphore organique et inorganique faiblement soluble, le phosphore inorganique insoluble, le phosphore fortement absorbé et le phosphore organique insoluble. Le phosphore disponible dans le sol est la fraction total de phosphore facilement accessible pour l’absorption des racines des plantes (Ziadi et al., 2013). En effet, moins de 1 % du phosphore total est assimilable par les plantes. Cette faible disponibilité du phosphore dans la solution du sol limite considérablement la nutrition phosphatée des plantes. Dans ces conditions, l’utilisation des phosphates naturels est à promouvoir, particulièrement en Afrique de l’Ouest, en raison de l’importance des gisements. Cependant, ce PN est peu assimilable par les plantes. Il a été démontré qu’il est mieux utilisé par les plantes agricoles en présence de mycorhizes à vésicules et à arbuscules. De plus, il est établi que la plante peut excréter des acides organiques qui augmentent la dissolution des PN dans le sol (Guissou et al, 1998; Ziadi et al, 2013).
Table des matières
1. INTRODUCTION
2. SYNTHESE BIBLIOGRAPHIQUE
2.1. Les symbioses mycorhiziennes
2.1.1. Généralités sur les symbioses mycorhiziennes
2.1.2. La symbiose mycorhizienne arbusculaire
2.1.2.1. Mise en place de la symbiose mycorhizienne à arbuscules
2.1.2.2. Développement de la symbiose mycorhizienne à arbuscules
2.1.3. La reproduction des champignons mycorhiziens à arbuscules
2.2. Importance des champignons mycorhiziens à arbuscules
Importance dans la nutrition des plantes
Importance face aux stress abiotiques et biotiques
Importance sur la structure du sol
2.3. Les phosphates naturels (PN)
3. MATERIELS ET METHODES
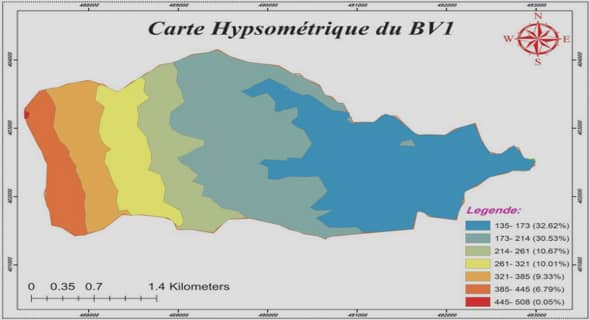
3.1. Présentation de la zone et du gisement d’étude
3.1.1. La zone d’étude
3.1.2. Le gisement d’étude : LAM-LAM
3.2. Echantillonnage du sol et des racines
3.3. Etude des champignons mycorhiziens à arbuscules (CMA)
3.3.1 Détermination du potentiel infectieux mycorhizogène
3.3.2 Extraction, dénombrement, morphotypage et identification des spores
3.3.3 Coloration des racines
3.3.4 Observations des CMA dans des racines colorées
3.4. Traitement des données
4. RESULTATS ET DISCUSSION
4.1. Mycorhization des racines des plantes récoltées
4.2. Potentiel mycorhizogène des sols (MPN)
4.3. Diversité et identification des spores de CMA
4.3.1 Diversité des spores
4.3.2. Identification des spores de CMA
4.3.2.1. Famille des Glomeraceae
4.3.2.2. Famille des Gigasporaceae
4.3.2.3. Famille des Acaulosporaceae
Conclusion et perspectives
REFERENCES BIBLIOGRAPHIQUES
WEBOGRAPHIE
ANNEXES