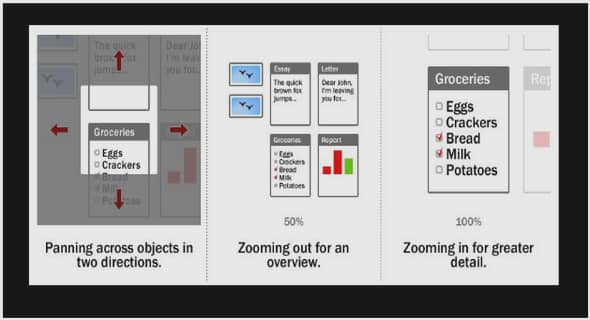
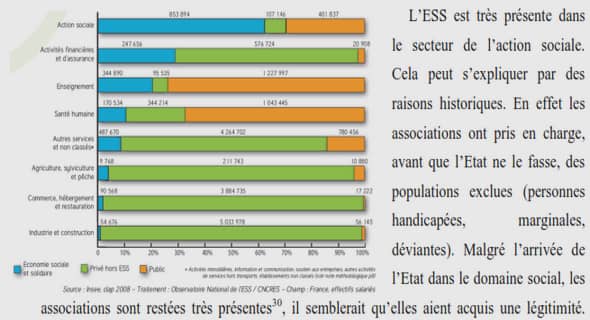
Bulletin des Biotechnologises – Mars 2002
Les gènes et les génomes
10. ### Les problèmes de duplication de gènes et de la polyploïdie ont encore suscité une revue. T Mitchell-Olds et al.; Current Opinion in Plant Biology 5 (FEB02) 74–79. On peut, en effet, aborder ce problème sur le plan fonctionnel avec les séquences complètes d’Arabidopsis et du riz. Des études ont récemment été consacrées à l’évolution des régulations, aux fonctions éventuelles des régions non codantes (ce qui est possible quand on dispose des séquences) notamment des séquences régulatrices, et l’effet de la sélection sur les fonctions des protéines. Une large fraction de la divergence des fonctions est certainement adaptative, et devrait permettre de comprendre les fonctions des gènes et des protéines codées. La revue résume les conclusions sur le problème de l’organisation des gènes les uns par rapport aux autres (synténies, notamment, ou association et ordre de gènes) Des synténies sont souvent observées entre espèces pour des marqueurs moléculaires, mais elles sont souvent masquées par des inversions, additions, délétions, duplications dans les séquences ADN. Pour les gènes ceci est moins gênant. Ainsi chez les Brassicacées, l’ordre des gènes est bien conservé d’Arabidopsis à Capsella et Brassica avec une distance évolutive de 10–14 et 16–21 millions d’années respectivement.
Chez les monocotylédones, les gènes présentent une bonne colinéarité, mais les régions intergéniques ont beaucoup évolué, avec, cependant, peu d’inversions et de duplications.
On avait déjà constaté que la localisation et les fonctions moléculaires des gènes de résistance au sein des Solanacées sont bien conservées chez la tomate, la pomme de terre, et le poivron, tout en laissant place à une évolution rapide de la spécificité. (voir RC Grube et al.; Genetics 155 (JUN00) 873-887). Ceci peut être utilisé, même avec des séquences partielles. On a ainsi, par extrapolation de microsynténies entre Arabidopsis et tomate par exemple, identifié dans un clone BAC (chromosome bactérien artificiel) le locus ovate de la tomate. On peut donc utiliser le génome d’Arabidopsis, et on a effectivement constaté une microsynténie, même entre riz et Arabidopsis portant sur cinq chromosomes et sur des ensembles allant jusqu’à 8 gènes colinéaires.
L’analyse des séquences régulatrices a été réalisée pour les promoteurs des gènes Chs, de la chalcone synthase, et Apetala3 (MA Koch et al.; Molecular Biology & Evolution 18 (OCT01) 1882-1891 chez une série de Crucifères. On a comparé la réponse à la lumière des différents promoteurs dans des protoplastes d’Arabidopsis. Elle est bien conservée. Ce qui est intéressant c’est que la fonction est conservée, même si la séquence est mal conservée. On l’avait déjà constaté chez la Drosophile. Il faut dire que c’est la fonction qui est sélectionnée, pas la séquence per se. Ceci va compliquer la détection des motifs régulateurs. Curieusement des séquences non transcrites et, pour l’instant, sans fonctions régulatrices, peuvent être conservées.
La fluidité du génome superposée à ces duplications emboîtées a été évaluée chez Arabidopsis. La demi-vie d’un gène individuel dupliqué est de 3,2 millions d’années, et la fréquence de duplication individuelle des gènes est de l’ordre de 0,02 à 0,002 par million d’années. Ce n’est pas négligeable. La survie de gènes dupliqués résulte probablement de la divergence des fonctions qui sont sélectionnées indépendamment. La divergence se traduit finalement par une barrière reproductive empêchant l’hybridation. On sait qu’elle existe et que plusieurs QTLs la gouvernent chez le tournesol, par exemple. La comparaison avec l’évolution parallèle des protéines de surface du spermatozoïde et leurs récepteur à la surface de l’ovocyte a été décrite pour certains animaux comme l’ormeau (Halyotis) où la lysine (qui disloque la barrière vitelline de l’ovule) montre une évolution rapide au sein d’espèces sympatriques (cohabitantes). Il y a sélection très forte de certains acides aminés de l’enzyme affectant la spécificité de reconnaissance du récepteur. On sait que la compétition interspécifique entre les grains de pollen joue un rôle de barrière à une hybridation interspécifique chez le tournesol et les iris.
Le développement
12. ### On trouvera dans NH Battey et al.; Current Opinion in Plant Biology 5 (FEB02) 62–68, une analyse du basculement du stade végétatif vers la floraison (floral transition). Il se trouve que bien des analogies peuvent être faites, au niveau moléculaire, avec une autre transition qui est celle du stade juvénile vers le stade adulte (acquisition de la compétence à fleurir). On commence à explorer ce phénomène chez un grand nombre de plantes annuelles où cela est plus facile pour essayer d’extrapoler vers les pérennes. L’identité végétative du méristème caulinaire de la plante est modifiée à la transition juvénile-adulte. Le méristème prend alors une identité d’inflorescence, puis florale. La notion d’identité résulte de l’existence des gènes homéotiques, c’est à dire de gènes entraînant l’apparition du méristème d’un organe donné à une place normalement occupée par un autre, cas de LEAFY (LFY) dont la mutation entraîne l’apparition de méristèmes d’inflorescence à la place d’un méristème floral. Mais on ne sait pas trop bien comment le sort du méristème est canalisé. On sait que LFYest indispensable au développement d’une fleur, mais la formation de la fleur n’est pas pré-programmée du fait de l’action de LFY sur le méristème lors de la transition florale. AGAMOUS (AG) est nécessaire à la formation des étamines et des carpelles au centre de la fleur, mais on ne sait pas ce qui limite son expression au centre de la fleur, alors que son activateur en amont, LFY est capable de le faire dans tout le méristème. On sait que le produit du gène WUS assure la persistance de la population méristématique, mais il coopère avec le produit de LFY dans l’activation d’AG. On a là une association d’un facteur organisateur du méristème et d’un facteur d’organisation de la fleur.