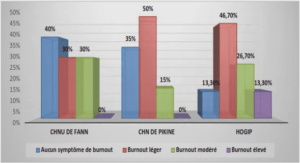
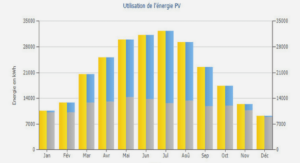
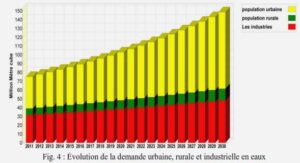
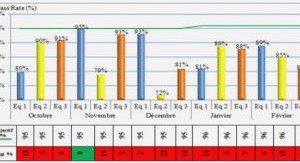
DETERMINATION DU POTENTIEL INFECTIEUX EN RHIZOBIUMS SYMBIOTIQUES DE MIMOSA PIGRA ET MIMOSA PUDICA DE SOLS
Dialogue Moléculaire
La reconnaissance mutuelle spécifique entre les deux partenaires symbiotiques se fait par un échange de signaux moléculaires ou dialogue moléculaire (Figure 1). Celui-ci permet l’infection et l’induction de l’organogénèse des nodules (Dénarié et Cullimore, 1993). Les rhizobia présents à l’état saprophytique dans la rhizosphère, répondent par un chimiotactisme positif aux exsudats racinaires (majoritairement flavonoïdes) de la plante, (Barbour et al., 1991). Les protéines NodD sont ainsi activées et induisent l’expression des gènes nod communs et spécifiques (Mulligan and Long, 1989 ; Fellay et al., 1995). Cette expression des gènes structuraux conduit à la production de signaux bactériens extracellulaires ou facteurs Nod qui sont responsables de la déformation des poils absorbants (Lerouge et al., 1990), de l’induction du méristème nodulaire et la formation des nodules chez la plante hôte (Dénarié et al., 1996 ; Long, 1996 ; Debellé et al., 2001 ; Parniske et Downie, 2003).
Formation de la nodosité: mécanismes d’infections
La formation des nodules ou nodosités sur les racines mais également sur les tiges des légumineuses a été bien étudiée (Vincent, 1980 ; Newcomb, 1981 ; Dreyfus et al., 1988 ; Sprent, 1989 ; Ndoye et al., 1994). Elle peut se faire de différentes manières suivant le mode d’infection. L’infection via les poils absorbants de la racine hôte est la plus répandue (Dart, 1977 ; Bauer, 1981 ; Verma et Long, 1983). Sous l’effet des facteurs Nod produits par la bactérie, les poils absorbants en croissance subissent des courbures caractéristiques dites «en crosse de berger» (Franssen et al., 1992). L’infection se fait par l’intermédiaire d’un cordon d’infection qui est une structure tubulaire d’origine végétale. Ce cordon se développe dans la lumière du poil, progresse vers la base et atteint les cellules du cortex racinaire, où il se ramifie (Turgeon et Bauer, 1985 ; Truchet et al., 1985, Pawlowski et Bisseling, 1996). Simultanément à l’infection, des cellules corticales se divisent activement pour former le primordium nodulaire dans lequel les bactéries vont se différencier en bactéroїdes capables de fixer l’azote et qui seront libérés dans les cellules de l’hôte (S mitt et al., 1992). Le primordium nodulaire, continue son développement pour former un nouvel organe végétal: le nodule (Dénarié et al., 1992). Dans d’autres cas, l’infection ne fait pas intervenir les poils absorbants et la pénétration des bactéries se fait, soit au niveau d’une zone de faiblesse de l’épiderme des racines latérales (crack entry) (Chandler, 1978 ; Dreyfus et al., 1988 ; James et al., 1992 ; Ndoye et al., 1994), soit à la jonction de deux cellules épidermiques latérales (infection intercellulaire) (de Faria et al., 1988). Il existe une grande variété de nodules qui diffèrent entre eux par leur anatomie, leur morphologie ou encore leur type de développement. Deux types principaux de nodules sont décrits : les nodules indéterminés de forme allongée et à activité méristématique continue qui se forment chez les légumineuses tempérées et les nodules déterminés qui sont sphériques à activité méristématique limitée dans le temps chez les légumineuses tropicales (Hirsch et al., 2001 ; Patriarca et al., 2002 )
Mécanisme de fixation de l’azote atmosphérique
L’azote est un élément majeur qui joue un rôle fondamental dans la constitution de la matière vivante. L’atmosphère qui est la principale réserve naturelle d’azote en contient 78% en volume. Cet azote moléculaire en abondance considérable est inaccessible à la plupart des Synthèse bibliographique 9 êtres vivants. Seuls des organismes appartenant aux groupes des procaryotes sont capables de le réduire sous une forme combinée assimilable (Boivin et Dénarie, 1998). Ce mécanisme appelé f ixation biologique de l’azote joue un rôle fondamental dans le cycle de l’azote et dans l’écosystème tout entier puisqu’elle constitue la plus grande source d’azote combiné produit dans la nature (Dreyfus, 1982). Il consiste en la réduction enzymatique de l’azote moléculaire (N2) en ammoniac (NH3) forme assimilable par la plante selon la réaction suivante: Mg++ N2 + 8H+ + 8e- + ATP ―→ 2NH3 + H2 + ADP + Pi Nitrogénase Cette réaction de fixation de N2 a un coût énergétique élevé (Gandy et Dommergue, 1995). Elle est catalysée par un complexe enzymatique réducteur: la nitrogénase (Witty et Minchin, 1988) qui est présente chez tous les microorganismes fixateurs d’azote (Blondeau, 1980 ; Kim et Rees, 1994 ; Rai et al., 2000). Très sensible à l’oxygène, la nitrogénase est protégée chez les légumineuses par la leghémoglobine qui contrôle le transport de l’oxygène et permet la respiration des bactéroïdes au sein du nodule (Halverson et Stacey, 1986). C’est à travers ce dernier que se font les échanges de substrats carbonés et azotés entre les deux partenaires symbiotiques.
La spécificité d’hôte
La spécificité d’hôte est une des caractéristiques majeures de la symbiose rhizobiumlégumineuses (Young et Johnston, 1989). C’est la capacité d’une souche à entrer en symbiose avec certains genres ou même certaines espèces de légumineuses. Cependant, elle s’applique aussi bien aux rhizobiums qu’aux plantes hôtes. La spécificité d’une souche peut être qualifiée d’étroite à plus ou moins large selon le nombre d’espèces de plantes hôtes. Un exemple classique de souche caractérisée par sa non-spécificité est Sinorhizobium sp., NGR 234 isolée de Lablab purpureus (Stanley et Cervantes, 1991) qui est capable d’établir une interaction symbiotique avec plus de 112 genres de légumineuses (Broughton et Perret, 1999). Contrairement à d’autres souches très spécifiques comme Azorhizobium caulinodans qui ne semble noduler efficacement qu’une seule espèce, Sesbania rostrata (Boivin et al., 1997 ; Boivin & Dénarié, 1998). Chez les légumineuses, les espèces du genre Galega ne peuvent Synthèse bibliographique 10 former de nodules qu’avec les souches de Rhizobium galegae (Lindström, 1989 ; Firmin et al., 1993; Terefework et al., 1998). A l’inverse, des espèces comme Macroptilium atropurpureum (Siratro) et Vigna unguiculata peuvent être noduler par différentes espèces de rhizobia (Somasegaran et Hoben, 1985 ; Lewin et al., 1987). La spécificité d’hôte est donc très variable. Elle est le fait d’un certain nombre de gènes dont les gènes de spécificité qui en sont les déterminants majeurs (Spaink, 1995 ; Dénarié et al., 1992). Cependant, il a été montré que les gènes communs nodA et nodC participent également à l a détermination du spectre d’hôte (Débellé et al., 1996 ; Roche et al., 1996 ; Kamst et al., 1997 ; Perret et al., 2000 ; Débellé et al., 2001). 2. Importance économique et écologique de la fixation biologique de l’azote La fixation biologique de l’azote est le processus biologique le plus important sur la terre après l’assimilation du ga z carbonique (CO2) durant la photosynthèse (Dixon et Wheeler, 1986). Elle est réalisée par deux groupes de microorganismes : les fixateurs libres et les fixateurs symbiotiques. Ces dernières sont les plus efficaces en termes de fixation car elles sont responsables de plus de 120 millions de tonnes d’azote atmosphérique par an (Elmerich et al., 1998). Les symbioses fixatrices d’azote (rhizobia-légumineuses, Frankia-plantes actinorhiziennes) jouent un r ôle écologique très important. En effet, les plantes impliquées dans ces symbioses acquièrent une certaine autonomie leur permettant de croître dans des sols pauvres, dégradés ou soumis à des stress (Date, 2000). Ainsi, la fixation biologique de l’azote permet la réhabilitation des sols dégradés et l’enrichissement minéral par l’intermédiaire de la litière (racines, nodules, parties aériennes) qui, après décomposition, libère l’azote sous des formes directement assimilables par les plantes (Rufty et al., 1983). L’installation d’essences ligneuses rendue possible grâce à la fixation biologique de l’azote a un rôle écologique important car elle permet la stabilisation et la colonisation des sols en le protégeant des érosions éoliennes et pluviales. D’un point de vue économique, la fixation biologique de l’azote est bien exploitée. En effet, elle améliore la productivité des plantes, favorisant ainsi une hausse des rendements agricoles. Elle est aussi utilisée dans les systèmes agroforestiers et dans les programmes de reboisement qui luttent contre la désertification et la déforestation.
INTRODUCTION |