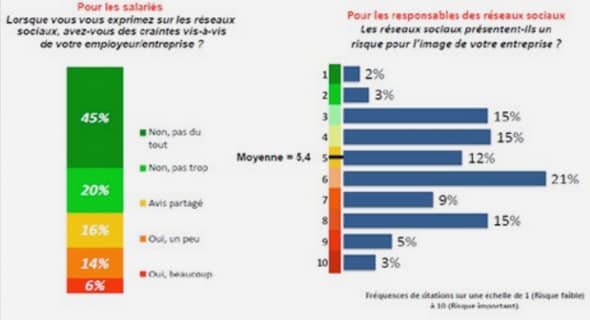
Répartition de la sauvagine au Québec
Le groupe d’espèces nommé « sauvagine » comprend des espèces de l’ordre des Ansériformes, la plupart faisant partie de la famille des Anatidea (canards, oies et cygnes) (Baldassarre et Bolden, 1994). Pour cette étude, nous nous intéressons également à certaines espèces d’oiseaux aquatiques faisant partie de l’ordre des Podicipediformes (comme les grèbes) et des Gaviiformes (comme les huards). Dans le cadre de cette étude, nous regroupons donc sous ce terme les différentes espèces de canards, d’oies, de cygnes, de grèbes et de huards En Amérique du Nord, les premiers efforts pour recenser les différentes espèces de sauvagine présentes sur le continent se sont manifestés dès les années 1960, avec différents programmes de baguage et par l’inventaire traditionnel de l’Ouest. Après avoir observé des déclins importants des populations de sauvagine, le Plan nord-américain sur la gestion de la sauvagine a été signé en 1986, dans l’objectif d’effectuer un plan de conservation des espèces qui serait cohérent sur l’ensemble du continent. Le suivi des espèces est depuis effectué par le moyen de survols aériens qui sont faits à l’échelle du continent. Pour l’Est du Canada, suite au délin observé des populations de Canard noir dans les aires d’hivernage, «Le plan conjoint sur le Canard noir» a été créé en 1990. Ce plan permet de fournir des indices sur les tendances populationnelles du Canard noir et des autres espèces de sauvagine nicheuses (Bordage et al., 2003). Parmi les données recueillies au cours des dernières années, il a été mis en évidence que 37 espèces de sauvagine utilisent régulièrement les plans d’eaux du Québec, que ce soit comme site d’hivernage, pour leur migration ou leur reproduction (Lepage et Bordage, 2013).
Au sein des forêts de la province, la densité d’individus serait relativement faible, soit moins de 1 couple nicheur/km2 selon Lemelin et al. (2004). Cependant, plus de 355 000 couples nicheurs ont été recensés dans le vaste milieu forestier du Québec (Bordage et al., 2003) et jusqu’à 19 espèces de sauvagine sont reconnues comme étant des nicheurs réguliers dans les forêts de la province. Ces aires supportent également 50% de la population mondiale du Canard noir (Mack et Morrison, 2006) et sont le coeur de l’aire de répartition de la population de l’Est du Garrot d’Islande (Robert et al., 2000), dont le statut a été désigné comme étant préoccupant par le Comité sur la situation des espèces en péril au Canada (COSEPAC, 2011). L’Ouest de la province serait également plus riche en couples nicheurs si on le compare au reste du Québec. Ainsi, en Abitibi-Témiscamingue, une moyenne de 1,5 couples nicheurs/km2 aurait été déterminée suite aux inventaires effectués dans les dernières années (Lemelin, 2004; Lemelin et al., 2008).
La superficie du plan d’eau et la présence d’eau libre
Plusieurs études effectuées sur l’utilisation de l’habitat ont démontré l’importance de la superficie d’un plan d’eau pour la sauvagine (Adde et al. 2020). Ainsi, McNicol et al. (1987) et Nummi et Pöysä (1995a) ont trouvé que les plans d’eau de petite superficie étaient plus utilisés par la sauvagine non-piscivore par rapport aux plans d’eau de plus grande taille. Lemelin et al. (2010) ont également confirmé cette relation en démontrant l’importance des plans d’eau de petite superficie (<8 ha) pour plusieurs espèces de sauvagine qui utilisent préférentiellement ces habitats. Différentes caractéristiques inhérentes à ces petits plans d’eau les rendent favorables à l’établissement des différentes espèces de sauvagine. Par exemple, l’exposition au vent et aux vagues est réduite dans les plans d’eau plus petits (Cowardin et al., 1979) et la faible profondeur des plans d’eau peut faciliter l’acquisition des ressources alimentaires par la sauvagine (Pöysä 1983). Lemelin et al. (2010) ont également démontré que la majorité des espèces utilisent très peu les eaux se situant loin des rives (soit à plus de 100m), ce qui pourrait indiquer que le nombre de couples dépend plutôt de la longueur du rivage disponible plutôt que de la superficie du plan d’eau. Les étangs, définis par Cowardin et al. (1979) comme «small, shallow, permanent or intermittent water bodies », seraient donc utilisés par plusieurs espèces, et parmi ces derniers, ceux présentant une certaine superficie d’eau libre (qui ne sont pas recouverts par une trop grande superficie de végétation) seraient d’une grande importance pour la sauvagine. Certaines études s’étant intéressées aux différents types de milieux humides utilisés par la sauvagine ont donc montré cette relation (Lemelin et al., 2010; Rempel et al., 1997), selon laquelle de nombreuses espèces sont associées avec les étangs semi-ouverts ou ouverts. Contrairement à ce qui a été trouvé pour les espèces de canards barboteurs et de canards plongeurs non-piscivores, les espèces consommant du poisson seraient plutôt associées avec les grands plans d’eau (Found et al., 2008; Lemelin et al., 2010), puisque la probabilité de retrouver du poisson serait plus grande dans ces derniers (McNicol et al., 1995).
L’abondance de ressources alimentaires
De façon générale, l’abondance des ressources alimentaires est un autre élément qui affecte l’utilisation de l’habitat par la sauvagine, et ce particulièrement pour les couvées au cours de la période de reproduction (Sugden, 1973). D’abord, les couvées auraient des besoins très spécifiques en ce qui a trait à leur alimentation. Plus particulièrement pour les espèces non-piscivores, les canetons se nourrissent surtout d’insectes durant les premiers jours et diversifient leur régime alimentaire au cours du temps (Collias et Collias, 1963). Il a aussi été montré que les femelles adultes s’alimentent d’invertébrés pendant la période de reproduction (Krapu, 1974; Reinecke et Owen, 1980). Dans les deux cas, il s’agit d’une période où la demande en protéines est très élevée pour la formation et la croissance des tissus animaux. Certains acides aminés sont dits essentiels, puisqu’ils ne peuvent pas être synthétisés par l’organisme et qu’ils doivent se retrouver dans les aliments consommés par un individu. Certaines espèces de sauvagine peuvent donc compléter leur alimentation en consommant des invertébrés aquatiques, qui sont très riches en acides aminés divers (Sugden, 1973). Les invertébrés aquatiques peuvent également être une source d’autres éléments essentiels, tels que certains acides-gras, qui sont synthétisés par les algues et qui seront accumulés par les invertébrés (Gladyshev et al., 2009; Grosbois et al., 2017; Schneider et al., 2017). De nombreuses études ont montré que les femelles quittaient l’étang de nidification une fois que les oeufs étaient éclos afin de se déplacer vers des étangs plus riches en ressources pour élever leurs couvées (Nummi et Pöysä, 1993; Ringelman et Longcore, 1982; Wayland et McNicol, 1994). Puisque les invertébrés sont une ressource essentielle pour les canetons à cette période, les femelles prioriseront donc des étangs où cette ressource est abondante. Ainsi, les facteurs agissant positivement ou négativement sur l’abondance d’invertébrés sont susceptibles d’affecter la sélection de l’habitat par la sauvagine.
Par exemple, plusieurs études ont montré que la présence de poissons dans un étang peut affecter l’abondance ou la richesse d’invertébrés aquatiques dans le milieu (Mallory et al., 1994; McNicol et al., 1995; McNicol et Wayland, 1992). En raison de la compétition existant entre la sauvagine et les poissons pour cette ressource, certaines espèces de sauvagine évitent les étangs occupés par les poissons et sont donc négativement associées avec leur présence (Epners et al., 2010; McNicol et Wayland, 1992) ou leur abondance (Eadie et Keast, 1982; Väänänen et al., 2012) dans le milieu. En revanche, puisque les espèces de sauvagine ont des modes d’alimentation qui diffèrent (barboteurs ou plongeurs) et qu’elles peuvent s’alimenter dans des types de milieux qui diffèrent également (en eaux libres ou dans des sections plus végétalisées des plans d’eau), les poissons n’ont pas le même effet sur toutes les espèces. Par exemple, le Garrot à oeil d’or est une espèce de plongeur qui s’alimente surtout en eau libre et qui consomme préférentiellement des invertébrés pélagiques (ou invertébrés de la colonne d’eau). Le Fuligule à collier (une autre espèce de plongeur), quant à lui, consomme surtout du benthos. Le fuligule est donc moins affecté par la présence de poissons que le garrot puisque les poissons seront plutôt présents dans l’eau libre et auront surtout un impact sur l’abondance d’invertébrés pélagiques qui sont des proies plus visibles (McAuley et Longcore, 1988; McNicol et Wayland, 1992; Väänänen et al., 2012).
En contre-partie, le garrot sera aussi en mesure de plonger pour consommer des organismes du benthos dans un milieu où ses proies préférentielles sont affectées, alors que les espèces de barboteurs seront généralement moins versatiles. Au sein du groupe des canards barboteurs, l’effet de la présence de poissons peut cependant varier entre les espèces. Par exemple, le Canard colvert consomme surtout des organismes du benthos et d’autres organismes présents sur la végétation. Des études ont ainsi montré que l’espèce s’alimente à proximité de la végétation, où il y a moins de poissons et sont donc peu affectés par ces derniers (Nummi et al., 2013; Väänänen et al., 2012). À l’opposé, ces mêmes études ont montré qu’une autre espèce, la Sarcelle d’hiver, qui s’alimente plutôt de proies en eaux libres, évite généralement les plans d’eau où il y a des poissons. Certaines études ont également montré un lien entre l’acidité du milieu, l’abondance de poissons et l’abondance d’invertébrés. Les poissons sont plus sensibles au changement d’acidité que les invertébrés en général, ce qui fait que sous un certain pH, les poissons ne peuvent pas subsister dans un étang donné alors que certains taxons d’invertébrés ne sont pas affectés. Pour certaines espèces de sauvagine, les étangs plus acides sont donc parfois préférés puisque la compétition pour les ressources invertébrées dans ces milieux est moindre (DesGranges et Darveau, 1985; McNicol et Wayland, 1992).
Cependant, plusieurs taxons d’invertébrés peuvent être affectés par le pH lorsque ce dernier est inférieur à 6, et de façon encore plus importante lorsque ce dernier passe sous le seuil de 5.5 (McNicol et al., 1995). Cela signifie que même si certains taxons d’invertébrés subsissent dans des étangs à pH plutôt faible, la richesse en invertébrés aura tendance à chuter de façon importante à partir d’un certain niveau La composition chimique du plan d’eau en phosphore total et en azote peut quant à elle avoir un impact sur l’abondance des ressources alimentaires puisqu’elle serait associée avec la productivité du milieu et son état d’eutrophisation (Nürnberg, 1996). Plus spécifiquement dans le contexte de la sauvagine, Pöysä et al. (2001) ont montré que la composition en phosphore total affecte l’utilisation de l’habitat par les couples avec des couvées, bien que cet indice à lui seul ne peut être utilisé pour déterminer quels plans d’eau seront surtout occupés par les différentes espèces. La productivité en nutriments et en macrophytes ont donc de l’importance puisque ces derniers permettent de soutenir les proies invertébrées dans le milieu (Longcore et al., 2006).
AVANT-PROPOS |