Généralités sur les Microsporidies et les Myxosporidies de Poissons
Les Microsporidies
Les Microsporidies ont longtemps été considérées comme des êtres pluricellulaires et classées à côté des Myxosporidies et Actinomyxidies dans le sous-embranchement des Cnidosporidies (POISSON, 1953). Il a fallu attendre l’essor de la microscopie électronique à transmission pour mettre en évidence leur nature unicellulaire et leurs caractéristiques très spéciales avec l’absence, notamment, de mitochondries typiques et de peroxysomes. Aujourd’hui les Microsporidies constituent un phylum à part dans le sous-règne des Protozoaires, celui des Microspora (SPRAGUE, 1977). Ce sont des parasites intracellulaires obligatoires. Elles s’attaquent à des représentants de presque tous les embranchements du règne animal depuis les protozoaires (VIVIER, 1975) jusqu’aux vertébrés les plus évolués y compris l’Homme (CANNING et LOM, 1986) ; chez ce dernier, prés de 10 espèces appartenant aux genres Encephalitozoon, Enterocytozoon, Septata, Nosema et Pleistophora ont été signalées, la plupart en relation avec le SIDA ou Syndrome de l’Immunodéficience Acquise (LIGIA et al., 2001). Certaines espèces peuvent être hyper parasites ; c’est le cas, par exemple, de Nosema marrionis hyperparasite de la Myxosporidie Ceratomyxa cori, elle- même parasite de la vésicule biliaire d’un poisson (CANNING et LOM, 1986). Les hôtes de prédilections restent cependant les arthropodes (principalement les insectes) chez les Invertébrés, et les poissons chez les Vertébrés.
Un poisson (l’Epinoche, Gasterosteus aculeatus) a d’ailleurs été le premier hôte chez lequel on a décelé la présence d’une Microsporidie (GLUGE, 1838). Cette Microsporidie a été nommée Nosema anomala par MONIEZ (1887), puis Glugea anomala par THELOHAN (1892). Depuis lors, les découvertes se sont succédées et, à ce jour, on compte une quinzaine de genres décrits dans diverses familles de poissons téléostéens dulçaquicoles ou marins à travers le monde (CANNING et LOM, 1986 ; FAYE et al. 1991 et 1996). Ainsi, plusieurs centaines de Microsporidies ont été répertoriées et classées dans les genres Glugea Thelohan, 1891 ; Thelohania Henneguy, 1892 ; Pleistophora Gurley, 1893 ; Ichthyosporidium Caullery et Mesnil, 1905 ; Marzekia Leger et Hesse, 1916 ; Enterocytezoon Desportes, Le Charpentier, Galian, Bernard, Cochand-Priollet, Lavergne, Ravisse, et Modigliani, 1985 ; Heterosporis Schupert, 1969 ; Nosemoides Vinckier, 1975 ; Spraguea Weissenberg, 1976 ; Tetramicra Matthews et Matthews, 1980 ; Loma Morrison et Sprague, 1981 ; Microgemma Ralphs et Matthews, 1986 ; Encephalitozoom Levaditi, Nicolau et Schoen, 1923 ; Microfilum Faye, Toguebaye et Bouix, 1991 ; Neonosemoides, Faye, Toguebaye et Bouix, 1996. A ces genres, il faut ajouter le groupe collectif Microsporidium Balbiani, 1884 qui renferme les Microsporidies en attente d’un statut systématique précis, leur cycle de développement étant encore insuffisamment caractérisé.
Cycle de développement
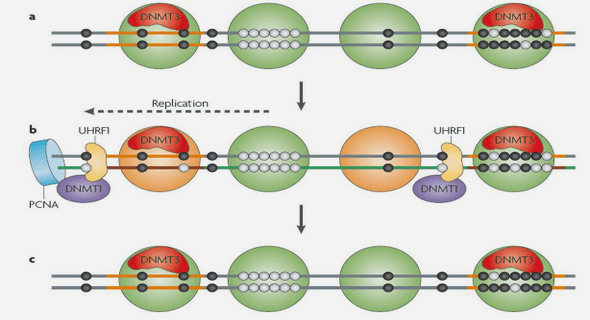
Les Microsporidies tirent l’énergie nécessaire à leur développement de la cellule hôte. Selon l’espèce, la multiplication (extrêmement rapide) aboutit à une infection localisée avec la formation d’une structure ‘’kystique’’ appelée xénome ou à une infection diffuse se traduisant par une invasion progressive du tissu atteint. Dans les deux cas, le développement reste semblable dans ses grandes lignes. Le cycle de développement des Microsporidies débute par l’inoculation du germe infectieux (sporoplasme) dans la cellule-hôte et s’achève par la formation des spores. Deux phases de développement sont connues : la mérogonie (ou schizogonie) qui est la phase de prolifération du parasite et la sporogonie, phase de maturation des spores (Fig. 1 et 2).
Le sporoplame expulsé de la spore peut se développer dans des tissus éloignés du site d’éclosion (tube digestif). En effet, BEKHTI (1984) a montré que les germes infectieux sont transportés par les polynucléaires neutrophiles jusqu’aux organes ou tissus cibles. Dans les cellules favorables, le sporoplasme se transforme en méronte qui s’accroît et se divise rapidement dans le cytoplasme de la cellule–hôte, augmentant ainsi le nombre de parasites. Pendant cette phase d’invasion de l’hôte, les mérontes et les plasmodes mérogoniaux constituent les stades de développement du parasite. Les mérontes peuvent présenter des noyaux isolés ou des diplocaryons, subir des divisions binaires répétées ou des divisions multiples. La dernière division du méronte donne naissance à des sporontes. Les sporoblastes subissent la maturation et deviennent des spores. Au cours de cette maturation, il y a synthèse des organites sporaux, augmentation de la quantité de réticulum endoplasmique lisse et granulaire ; les vésicules golgiennes sont responsables, en partie, de la sécrétion du filament et du sac polaires ; un endoplasme transparent est graduellement ajouté à la paroi sporale sous la couche dense maintenant appelée exospore. A la fin de la maturation, les vésicules atrophiées de l’appareil de Golgi fusionnent pour former la vacuole postérieure.