Des structures particulières
Les racines sont généralement classées de manière assez approximative en fonction de leurs diamètres (fines, moyennes, grossières, etc…) ou de leur localisation dans le sol (superficielles, peu profondes et profondes). Ces classifications ne sont pas exactement fondées sur une justification fonctionnelle ou physiologique (Zobel 2005). Le classement général de racines en fonction de leur diamètre est empiriquement (et approximativement) lié à leurs différences fonctionnelles. Les plus grosses racines sont généralement considérées comme des vecteurs de transfert de l’eau à longue distance entre le système racinaire et les parties aériennes de la plante. Les racines fines sont plus spécifiquement dédiées à des fonctions locales, telles que l’absorption d’eau et de nutriments. Cependant, comme le souligne Zobel (2005), la classification des racines fines comme rapportée dans la littérature par Großkopf (1950) ou Kreutzer (1961) manque de cohérence en ce qui concerne les propriétés physiologiques et la variabilité des diamètres. La plupart des racines sont généralement considérées comme «grossières» lorsque leur diamètre est >2 mm ou parfois > 5 mm, mais la justification d’un tel seuil reste floue et n’est pas consensuelle.
Les structures et fonctions des racines peuvent varier dans les différentes parties du système racinaire d’une même plante, mais cette variabilité dépend probablement du type fonctionnel de l’ensemble du système racinaire étudié. Par exemple, les racines latérales peu profondes du pin possèdent des cellules du xylème à paroi mince avec de grandes lumières, favorisant l’absorption rapide de l’eau. Pour la même espèce, les cellules du xylème des racines verticales sont à paroi épaisse avec de petites lumières (Marciszewska and Tulik 2013). Dans ce cas, l’absorption d’eau est moins efficace, mais la racine elle-même est plus résistante mécaniquement au poids de l’arbre situé directement au-dessus (Chaffey 2013; Fayle 1968). La structure des tissus végétaux et le diamètre des vaisseaux, diffèrent entre les tiges, les racines de surfaces et les racines profondes (Figure 4). Plus les racines sont profondes, plus le diamètre des vaisseaux est large (McElrone et al., 2004). Cette même étude (McElrone et al., 2004) a également mesuré l’anatomie du xylème, ainsi que la vulnérabilité à la cavitation et la conductivité hydraulique spécifique, pour quatre espèces d’arbres à croissance lente et résistantes à la sécheresse (Juniperus ashei , Bumelia lanuginosa , Quercus fusiformis et Quercus sinuata).
Ces recherches ont montré que les racines recueillies dans des grottes jusqu’à 20 m de profondeur diffèrent sensiblement de leurs homologues moins profondes. Avec de plus grands vaisseaux du bois et une conductivité hydraulique plus élevée, les racines profondes réduisent la résistance à l’écoulement et l’absorption d’eau profonde est maximisée. Mais cela se fait au prix d’une plus grande vulnérabilité à la cavitation. Le débit d’eau à travers le xylème des racines ligneuses est généralement sans restriction dans les deux sens de la redistribution (Schulte 2006). Des observations de l’ultrastructure des racines de la forêt boréale de conifères d’espèces Pinacée révèlent que la vulnérabilité à la cavitation est liée à l’épaisseur des membranes et à la position en profondeur des racines (Hacke and Jansen 2009). Néanmoins, plus de travail est nécessaire pour évaluer si ces corrélations existent dans d’autres groupes taxonomiques et s’ils varient selon le type de racines et/ou leur position dans le système racinaire. La combinaison de mesures expérimentales des caractéristiques hydrauliques des racines de maïs ( Zea mays L.) avec la distribution des racines simulées a montré qu’à une profondeur de 1 m, 80% des racines latérales du maïs ont atteint leur capacité maximale de transport d’eau au bout de 100 jours (Pierret et al. 2006).
Cependant, cette capacité maximale n’a été atteinte que dans 10 % des racines les plus profondes, responsables du transfert d’eau à longue distance à partir du point d’origine des racines. Watt et al. (2008) ont également mis en évidence que la plupart des racines les plus profondes des céréales ont un faible débit d’eau axial. Si l’existence de ces points de résistance majeurs au transfert d’eau à longue distance est une règle générale chez les espèces herbacées, ce phénomène pourrait représenter une limitation importante de la contribution des racines profondes à l’absorption de l’eau. Rewald et al. ont décrit en 2011 des fonctions physiologiques différentes en fonction des ordres des racines ayant des densités de tissus variables chez Citrus volkameriana. Bien que les racines hautement conductrices soient impliquées dans les interactions entre la végétation et les eaux souterraines profondes, il est peu probable que ce soient les organes d’absorption à travers lesquels l’eau du sol est extraite en profondeur. Au contraire, comme l’a suggéré McElrone et al. (2004), de vastes réseaux de racines fines absorbantes doivent surement exister en amont de ces collecteurs profonds. L’analyse récente de Lobet et al. (2014) décrit les mécanismes racinaires contribuant à l’extraction de l’eau en zone sèche et met en avant les possibilités d’extraction d’eau profonde pouvant être mises à profit dans des conditions agronomiques spécifiques.
Fonction des racines et rôles dans les processus de l’écosystème
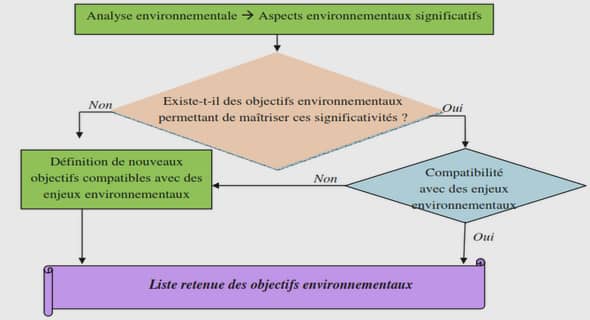
Les échanges et interactions liés au développement des racines profondes se trouvent au centre de nombreux mécanismes et ont un impact à plusieurs niveaux de l’écosystème terrestre. La Figure 5 ci-dessous schématise ces phénomènes, au travers des principaux rôles et fonctions liés aux racines.
a) Impacts des racines sur la physique du sol Les racines peuvent être considérées comme des ingénieurs du sol au travers de leurs différentes fonctions et rôles (Figure 5). Certains effets de la croissance des racines et de leur développement sur les propriétés physiques du sol sont bien documentés, en particulier dans l’étude proposée par Angers and Caron (1998) et résumés plus récemment par Pierret et al. (2011). L’un des changements les plus importants dans la structure du sol lié à la croissance des racines est la création de macropores continus. De tels macropores sont souvent associés à la pénétration active de racines dans les couches du sol de densité apparente élevée. La capacité des racines à perforer le sol varie en fonction des espèces (Bengough et al. 1997; Pierret et al. 2011a; Pierret et al. 1999) avec par exemple un plus grand potentiel de pénétration pour les dicotylédones par rapport aux monocotylédones. (Materechera et al. 1991) La présence des racines et des canaux radiculaires augmente l’infiltration de l’eau dans le sol. Les racines grossières dans les horizons plus profonds du sol favorisent l’infiltration de l’eau et réduisent le risque de ruissellement induit par la saturation des horizons peu profonds lors de pluies de fortes intensités (Jost et al., 2012). La croissance des racines dans le sol structuré ou compacté peut influencer la conductivité hydraulique du sol (Nobel and Cui, 2010). Les conséquences d’une infiltration accrue sont diverses pour la structure du sol, sa stabilité, les procédés chimiques et de l’activité microbienne (Ghestem et al., 2011) et ont en partie été modélisées (Dupuy et al., 2005a; Dupuy et al., 2007; Tobin et al., 2007). Les arbres à racines profondes réduisent les écoulements latéraux de subsurface et favorisent l’absorption d’azote dans les couches profondes du sol, ce qui représente un moyen efficace et durable de réduire les pollutions diffuses (Wang et al., 2011). L’architecture des racines contribue à la résistance au cisaillement du sol (Fan and Chen, 2010).
b) La microbiologie du sol Les racines des plantes ont une plasticité leur permettant de répondre rapidement aux changements de l’environnement local. Cette plasticité dépend en partie de la présence de microorganismes du sol, tels que les bactéries et les mycorhizes (Hodge et al. 2009). Il existe une association mesurable entre le nombre de bactéries et la densité d’enracinement, et par conséquent, un impact potentiel de la profondeur d’enracinement sur la propagation et la survie des communautés microbiennes du sol (Pankhurst et al. 2002; Pierret et al. 1999; Troxler et al. 2012). En se basant sur la description de Hiltner en 1904 et la définition proposée par Hinsinger en 1998, il n’y a aucune raison pour que l’on ne puisse pas qualifier le sol autour des racines profondes de « sol rhizosphérique ». Pourtant peu d’études abordent ce thème de recherche. Le sol inclus dans la rhizosphère a des propriétés sensiblement différentes de celles du reste du volume du sol, en raison principalement de l’exsudation racinaire qui affecte localement les teneurs en eau (Carminati and Vetterlein 2012) et éléments nutritifs (Hinsinger et al. 2009). En profondeur la rhizosphère joue un rôle important. Markewitz et al. (2010) indiquent qu’environ 40 % des précipitations terrestres totales transitent dans le « faible» volume de sol de la rhizosphère autour des racines des plantes, avant d’être transpiré, ce qui en fait une des régions les plus actives hydrauliquement de la biosphère (Bengough 2012). L’action physique des racines, en conjonction avec l’activité biologique dans la rhizosphère et les spécificités physico-chimiques qu’elle induit, permettent d’établir des canaux relativement stables, ou macropores (Ghestem et al. 2011). Ces canaux peuvent être riches en nutriments (Watt et al. 2008) et fournir un milieu favorable pour les micro-organismes (Pankhurst et al. 2002). De nouvelles racines peuvent se développer à travers ces macrospores, surtout dans les sols résistants à la pénétration, qui sont souvent présents dans les couches plus profondes du sol (White and Kirkegaard 2010).
Remerciements / Acknowledgements |