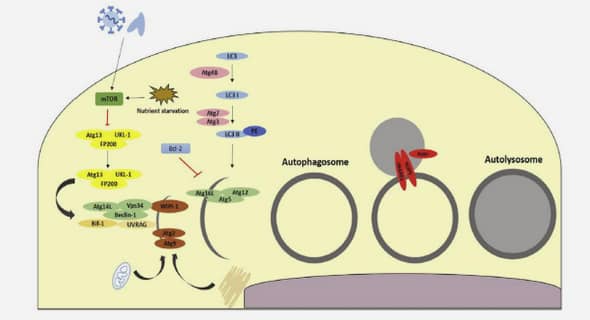
Mécanismes pathologiques de Nosema ceranae chez l’abeille
Présentation
Après la détection de N. ceranae sur A. mellifera en 2005, et avec la diminution de la population d’abeilles au niveau mondial, les efforts de recherche ont été orientés principalement vers l’étude de la prévalence et de l’épidémiologie de N. ceranae dans différentes régions du monde. Egalement de nombreux travaux ont été focalisés sur l’étude du développement du parasite dans l’intestin de l’abeille au niveau histologique, de la multiplication et la production des spores dans différentes conditions et de la mortalité des abeilles. Un investissement important a été fait aussi dans la mise au point de méthodes de diagnostics microscopiques mais surtout moléculaires pour la détection de N. ceranae et sa différenciation de N. apis. Cependant, la connaissance des mécanismes de base de l’infection notamment au niveau moléculaire, reste assez limitée avec des travaux sur l’activation du système immunitaire et les niveaux de tolérance à l’infection (e.g. Antúnez et al., 2009; Bourgeois et al., 2012 ; Chaimanee et al., 2012 ; Huang et al., 2012). Basés sur des pistes des recherches issues des études sur les interactions hôte – parasite chez d’autres modèles biologiques, insectes et mammifères, nous avons développé une hypothèse spécifique sur les mécanismes d’infection et la réponse de l’abeille à N. ceranae. Cette hypothèse considère que si l’épithélium de l’intestin agit comme une première ligne de défense face à une infection, comme le cas de N. ceranae, on pourrait s’attendre à l’activation de voies métaboliques du système immunitaire et la production d’espèces réactives de l’oxygène d’action antibactérienne. Dans le chapitre 3 nous développons ainsi notre deuxième objectif qui est l’exploration des mécanismes à la base des effets de N. ceranae. Cela par l’observation des différences entre des intestins d’abeilles infectées et d’abeilles saines en utilisant des méthodes moléculaires (transcriptomique) et biochimiques.
Nosema ceranae induces oxidative stress in the midgut
We used a tiling microarray based on gene predictions and annotation from the honey bee genome sequencing project [27] to identify at the gut level the pathological impact of N. ceranae on worker bees at seven days post-infection. A total of 9,293 genes were expressed in the gut, which represents approximately 70% of the genes so far identified from the honey bee genome. Out of those genes, the transcription of 336 genes was found to be significantly altered by the proliferation of N. ceranae spores (see Table S1in “Annexe 3.2” for the list of genes). Since we analyzed bees from 3 different colonies (i.e. different genetic backgrounds) that were each headed by a multi-drone inseminated queen, those genes might represent the ‘‘general’’ gut pathology and/or response to the parasite. When challenged by microorganism infections, insects can rapidly mount a potent immune response involving different molecular pathways [31]. For example, ingestion of bacteria activates the Imd pathway in the gut [32]. An activation of the innate immune system was therefore expected after challenging the bees with microsporidia, as it was found in drones [39]. However, no immune gene was more highly transcribed in workers after a seven-day infection. On the contrary, 2 genes (basket (GB16401) and u-shaped (GB16457)) that are involved in Drosophila immune responses [40,41] were downregulated (Table S1 in “Annexe 3.2”), perhaps indicating an immune suppression by the parasites, as suggested by Antúnez et al. [9]. Several studies have shown that one of the most immediate immune response of the gut involves the production of reactive oxygen species (ROS) to fight bacterial infection both in mammals [30,42] and insects [34,35,43]. ROS, which are efficient antimicrobial molecules, are generally derived from oxidation-reduction process. We performed a Gene Ontology analysis to explore which functional components were affected by N. ceranae, and found that genes involved in ‘‘oxidation reduction’’ were significantly overrepresented in the gene set upregulated upon spore infection (Table 1). This increase of oxidation reduction in the gut epithelia of bees parasitized by N. ceranae would therefore indicate an enhanced generation of ROS in response to the infection and suggests that ROS production is a general gut immune response to microorganism infection, including microsporidia. Since residual ROS can cause inflammatory disease, a balance between synthesis and elimination of ROS via antioxidants is necessary to protect the host gut [29,34,35,43]. Therefore, the antioxidant system may play an essential role during gut infection. Interestingly, the functional analysis also revealed a ‘‘response to oxidative stress’’ in the bee gut (Table 1, see Table S2 in “Annexe 3.3” for the list of genes associated with each Gene Ontology category), notably with the upregulation of the catalase (GB30227) and glutathione peroxidaselike 2 (Gtpx2 (GB18955)) genes, which have both antioxidant properties. A similar catalase, the immune-regulated catalase has been shown to be a key player of the Drosophila defense system during microbe infection in the gut epithelia [35]. Two cytochromes P450 were induced in the bee gut (CYP6AS4 (GB15793) and CYP6BC1 (GB10466)). CYP6AS4 has been shown to metabolize quercetin contained in honey and pollen [44], but it is not known whether they might contribute to the production or elimination of ROS. We further tested whether the protective response in the bee gut increased in response to N. ceranae infection by measuring the activity of major antioxidant and detoxification enzymes: superoxide dismutase (SOD), glutathione peroxydase (GP), glutathione reductase (GR) and glutathione-S-transferase (GST). The activity of SOD and GR was not significantly different between control and infected bees (Mann-Whitney U tests: p = 0.931 and p = 0.558, respectively; Fig. 2). Unexpectedly, the general activity of GP was significantly decreased by the spore infection (p = 0.002). We observed the opposite pattern for Gtpx2 at the transcriptome level (see above). The lower GP activity may either reflect a post-translational modification or the expression level of the other GP found in the bee genome (Gtpx1 (GB14138)) but its expression was not significantly different. The gut might also respond to a potential inhibition of GP activity upon spore infection by increasing the transcription of Gtpx2. Further experiments are needed to explore the GP pattern. Finally, the GST activity was induced by N. ceranae (p = 0.04 Fig. 2), as recently found by Vidau et al.
Nosema ceranae impairs cell signalling and tissue integrity in the midgut
A significant number of genes involved in cell signaling (e.g. of GO terms: ‘‘positive regulation of cell communication’’, ‘‘enzyme linked receptor protein signaling pathways’’, ‘‘transmembrane receptor protein tyrosine kinase signaling pathway’’) was inhibited by the parasites (Table 1). Cell-cell communication enables cells to perceive and correctly respond to their environment during tissue development and repair or the regulation of tissue homeostasis. Accordingly, the function ‘‘tissue homeostasis’’ was downregulated in parasitized bees, as well as biological processes implicated in ‘‘morphogenesis of an epithelium’’ and ‘‘open tracheal system development’’ (Table 1). The deregulation of ‘‘protein amino acid phosphorylation’’ by the parasite (Table 1) might be involved in the degeneration of the gut tissue. Since protein phosphorylation regulates many aspects of cell life, the modification of the phosphorylation states of intracellular proteins might be a cause or a consequence of the disease state [46]. These results suggest that the proliferation of N. ceranae caused a degeneration of the gut epithelium. Interestingly, the effect of N. ceranae on tissue morphogenesis and integrity at the molecular level was confirmed at the histological level (Fig. 3) as in a previous study [2]. The epithelial cells of infected bees showed major signs of degeneration, which are linked to the downregulation of biological process like ‘‘positive regulation of cell communication’’ and ‘‘tissue homeostasis and morphogenesis’’ (Table 1). Gut cells are usually renewed via the multiplication and differentiation of stem cells in the basal cell layer that, once differentiated, move toward the lumen. In insects, this renewal of intestinal stem cells is controlled by the canonical Wnt signaling pathway, which includes the main downstream components frizzled and armadillo [47,48]. Our results indicated that four main genes (frizzled2 (GB12765), groucho (GB13456), basket (GB16401) and armadillo (GB12463)) from this pathway were inhibited by the parasite, suggesting that N. ceranae development inhibited the self-renewal of intestinal cells of the host. This finding might be surprising, since the reaction of the gut to microorganisms involved not only the activation of the immune system, but also integrated responses controlling self-renewal and differentiation of stem cells, essential to the gut tissue homeostasis [32]. However, similar results have been found with Encephalitozoon microsporidia that induces a disruption of the cell cycle of the host cells [49] but without killing the host. The dramatic lifespan reduction of bees parasitized by N. ceranae could likely be explained by greater changes in the host cell cycle as compared to changes induced by non-lethal microsporidian species.