Evaluation des réponses en sous-classes d’IgG
anti-AMA1 et anti-MSP1p19 dans le paludisme
Paludisme simple
Il s’agit d’un paludisme symptomatique associé à une parasitémie sans signes de gravité ou de dysfonctionnement d’un organe vital (16). La primo-infection se caractérise par une phase d’incubation asymptomatique d’une durée variable pouvant aller de quelques jours à quelques mois. Apparaissent ensuite les premiers signes, peu évocateurs, de la maladie qui coïncident avec la multiplication des parasites dans le sang. On observe alors une fièvre modérée (38°-38.5°C), sans périodicité particulière, parfois accompagnée de maux de tête, de douleurs musculaires, de troubles digestifs, de vomissements, de diarrhées, et d’un affaiblissement généralisé. La maladie évolue ensuite vers une période d’état, qui, sans traitement spécifique, se caractérise par la succession d’accès palustres marqués par une phase de frissons intenses (17), une phase d’hyperthermie, et enfin, une phase de sueurs annonçant la fin de la crise et s’accompagnant d’une sensation de bien-être. On observe parfois une splénomégalie associée à de l’anémie. Page | 8 Ces accès fébriles correspondent à l’éclatement des globules rouges parasités. Leurs rythmes dépendent de l’espèce parasitaire impliquée et de la durée de son cycle : 2 jours pour P. falciparum ou P. vivax (fièvre tierce), 3 pour P. malariae (fièvre quarte)
Paludisme grave
Il s’agit souvent du paludisme grave à P. falciparum associé à des signes de gravité ou des signes de dysfonctionnement d’un organe vital (Tableau I). Tableau I: Critères de gravité du paludisme (16) Critères cliniques Altération de la conscience ou coma aréactif : Echelle de coma de Glasgow < 11 pour les adultes Echelle de coma de Blantyre < 3 pour les enfants Prostration : Faiblesse généralisée empêchant le patient de marcher ou de s’asseoir sans assistance Convulsions multiples : Plus de deux épisodes en 24 heures Respiration profonde, détresse respiratoire : Respiration acidosique Collapsus cardiovasculaire ou état de choc Tension artérielle systolique < 70 mm Hg chez les adultes < 50 mm Hg chez les enfants Ictère avec signes de dysfonctionnement d’autres organes vitaux Saignements spontanés anormaux Œdème pulmonaire : Respiration rapide avec râles bilatéraux aux bases pulmonaires Critères biologiques Hypoglycémie : Glycémie < 2,2 mmol/litre ou < 40 mg/dl Acidose métabolique : Bicarbonates plasmatiques < 15 mmol/litre Anémie normocytaire sévère : Hémoglobine < 5 g/dl, hématocrite < 15 % Hémoglobinurie Hyperparasitémie : Parasitémie > 4% chez le sujet non immun Hyperlactatémie : Acide lactique > 5 mmol/litre Insuffisance rénale aiguë : Créatinine sérique de 265 mmol/litre ou davantage Page | 9 On considère qu’un patient est atteint de paludisme grave à P. falciparum lorsqu’on décèle la présence de formes asexuées de P . falciparum dans son sang associée à une manifestation clinique ou biologique du paludisme sévère définie par l’OMS (Tableau I). P. vivax et récemment P. knowlesi, sont deux causes reconnues de formes palustres graves, essentiellement en Asie et dans certaines régions boisées d’Asie du Sud-Est. Le paludisme grave à P. vivax peut donner lieu à des manifestations cliniques analogues à celles du paludisme grave à P. falciparum et peut être fatal (16).
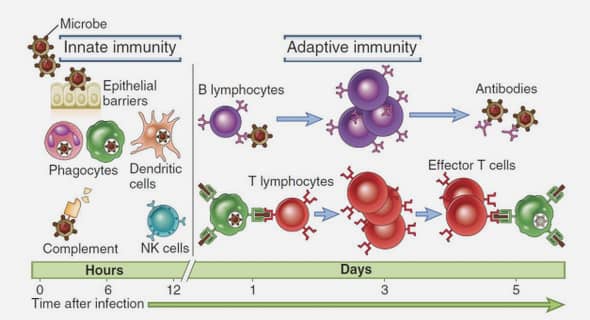
IMMUNITE ANTI-PALUSTRE
Elle se développe graduellement après une longue exposition aux infections plasmodiales à répétition dans les zones où le niveau de la transmission est élevé (zone stable). L’immunité stérilisante semble très rare puisque la prévalence parasitaire chez l’adulte approche 100% dans les zones endémiques (18). Les mécanismes exacts soutenant l’acquisition de l’immunité au paludisme sont mal compris, mais il est clair que les réponses innées jouent un rôle important (19). V.1. Notion de résistance naturelle A l’inverse des personnes souffrant de paludisme aggravé, certains individus sont moins sensibles, voire même réfractaires, à l’infection primaire. Cela peut s’expliquer par : Certaines anomalies génétiques altérant le développement du parasite dans le globule rouge. Il s’agit de : Hémoglobinopathies : Ce sont des types hémoglobiniques capable d’inhiber la croissance intracellulaire de P. falciparum. − L’ Hb S semble protéger les paludéens contre de fortes parasitémies (20). − L’ Hb F inhiberait la croissance de P. falciparum, ce qui expliquerait en partie, la rareté des accès palustres chez le nourrisson (21). − Les désordres quantitatifs portant sur une chaîne hémoglobinique (thalassémie) peuvent aussi ralentir la croissance du Plasmodium (20). La déficience en G6PD déshydrogénase aurait aussi un rôle protecteur (22) Page | 10 L’absence de molécules de surface du globule rouge indispensables à la pénétration du parasite, telles que : La molécule du groupe Duffy pour P. vivax et P. knowlesi (23,24). La glycophorine : A une glycoprotéine de membrane des globules rouges humains pour P. falciparum (25). I1 existe probablement d’autres mécanismes naturels de protection contre les stades érythrocytaires mais aussi exo-érythrocytaires du Plasmodium. Cependant, l’infection par Plasmodium engendre chez l’hôte des réponses immunitaires. Il s’agit de réponses non spécifiques dites innées et spécifiques ou acquises.
Réponse immunité innée
Parvenus dans l’organisme de l’hôte, les parasites peuvent pénétrer certains types de cellules immunitaires dans lesquelles ils se développent. Les mécanismes innés de l’inhibition de la croissance des parasites par l’hôte humain seraient probablement la cause du faible taux de parasitémie observé au cours des infections aigues à P. falciparum
Reconnaissance du parasite
Plusieurs récepteurs exprimés par ces cellules immunocompétentes seraient impliqués dans la reconnaissance de produits de P. falciparum. V.2.2. Rôle des cellules immunitaires Parmi les cellules en première ligne dans la réponse immunitaire innée après infection par P. falciparum, on distingue les cellules dendritiques (DC), les lymphocytes Tγδ et les macrophages. Les DC produisent de l’IFN-α en réponse à des lysats d’hématies infectées par P. falciparum (28). Cette activation serait importante pour aider à la production d’IFN-γ par les cellules Tγδ (28). Les cellules Tγδ sont aussi capables de reconnaître des molécules produites par P. falciparum et de produire de l’IFNγ. Ces cellules cytotoxiques contre des GRp par P. falciparum (29,30). Néanmoins, leur activation semble dépendre de cytokines exogènes, Page | 11 suggérant ainsi que les réponses Tγδ pourraient être aussi dépendantes de l’activation par d’autres types cellulaires, comme les monocytes et les cellules dendritiques (29). Tableau II: Molécules intervenant dans la reconnaissance de P. falciparum par les cellules du système immunitaire innée (27). Les monocytes/macrophages et les polynucléaires neutrophiles humains seraient également activés. Ils interviennent dans la phagocytose directe et dans la production de cytokines pro-inflammatoires comme le TNFα, l’IL1, le GM-CSF, l’IL18, des dérivés toxiques de l’oxygène et de l’azote responsables de la destruction du parasite au stade intrahépatique. Les cellules NK sembleraient jouer un rôle prépondérant dans l’immunité innée au cours des infections palustres précoces. Leur nombre augmente particulièrement et elles sont capables de détruire les GRp par P. falciparum (31,32). Elles sont aussi de puissantes productrices de cytokines telles que l’IFNγ qui conduit à l’activation parasiticide des macrophages et à l’augmentation de leur potentialité à détruire les globules parasités de l’hôte .Les cellules dites NK-T apparentés aux cellules NK sont de puissants inhibiteurs de la réplication parasitaire du stade hépatique dans les systèmes in vitro du paludisme de la souris (34). V.3. Immunité adaptative Elle est acquise progressivement avec le temps et intervient par des mécanismes effecteurs dirigés contre les différents stades évolutifs du parasite (mérozoïtes, schizontes, gametes et sporozoïtes). L’immunité acquise anti- plasmodium est associée aux faibles taux de parasitémies et aux épisodes cliniques de la maladie (35,36). Dans cette partie nous n’aborderons que l’immunité dirigée contre les stades sanguins asexués. Elle se divise en réponse humorale et cellulaire.
Réponses cellulaires
Les mérozoïtes libres et les hématies infectées, n’exprimant pas à leurs surfaces de molécules HLA, ne sont souvent considérés comme cibles des réponses immunes protectrices à médiation cellulaire. Cependant le rôle de l’immunité cellulaire dans la protection contre le paludisme a été démonstré par Pombo et al (37). Dans son étude, les personnes protégées présentaient de fortes proliférations cellulaires T CD4+ et T CD8+, une réponse cytokinique composée d’IFNγ, mais une absence d’interleukine 4 ou d’interleukine 10 et la production de fortes concentrations de NO dans les cellules mononuclées du sang des sujets. Les réponses immunitaires cellulaires débuteraient ainsi par l’activation des cellules T CD4+ dans la rate, après la présentation de l’Ag par les Cellules dendritiques. L’immunité cellulaire T serait régulée par l’IL-12 et ferait intervenir l’IFNγ et le TNFα qui, induisent la phagocytose des GRp ainsi que la mort des parasites intracellulaires via la synthèse de NO et des radicaux oxygénés par les neutrophiles et les macrophages. La destruction des parasites a lieu essentiellement dans la rate.
PREMIERE PARTIE : GENERALITES SUR LE PALUDISME |