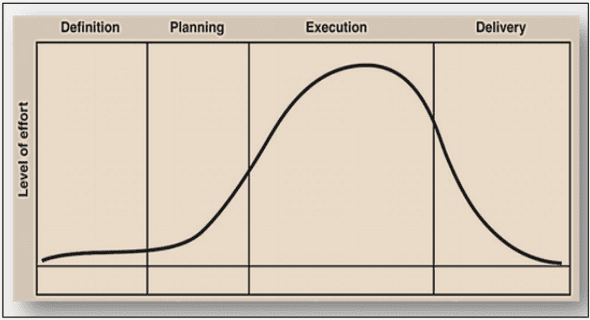
La traduction eucaryote entre biologie et physique
Introduction au processus de traduction
La traduction est un processus cellulaire commun à l’ensemble des règnes du vivant, qui correspond au processus de fabrication des protéines à partir du code génétique. Dans les cellules procaryotes ou eucaryotes, l’information génétique est portée par l’acide désoxyribonucléique ou ADN. Cette macromolécule double-brin contient l’ensemble de l’information génétique, aussi appelé génome, d’un être vivant. L’ADN est composé de 4 bases nucléiques : l’adénine (A), la cytosine (C), la guanine (G) et la thymine (T). Les molécules d’ADN sont stables dans le temps, ce qui assure la pérennité du code génétique, mais elles ne peuvent être directement transformées en protéines. Une enzyme appelée ARN polymérase va réaliser une copie d’une partie de l’ADN. Cette étape nommée transcription aboutit à la formation de l’ARN messager (ARNm). Cette copie de l’ADN, qui se dégrade rapidement dans le temps, est une macromolécule simple brin, ellemême composée de 4 sortes de bases nucléiques : l’uracile (U), la cytosine (C), la guanine (G) et l’adénine (A). Après des étapes de maturation qui ont lieu pour les eucaryotes dans le noyau cellulaire, l’ARN messager est exporté dans le cytoplasme pour y être traduit en protéine. On appelle le processus de production des protéines traduction car il correspond au passage de l’alphabet de l’ARN messager (les bases nucléiques A,U,G,C), à l’alphabet des protéines composées d’acides aminés (il en existe 22 sortes différentes). La macromolécule responsable de la traduction est le ribosome. Le ribosome lit l’ARN messager codon par codon, un codon, groupe de trois bases nucléiques, correspondant à un acide aminé de la chaine protéique. Il assemble au fur et à mesure la chaine protéique et la relâche lorsqu’elle est complète. Le ribosome est capable de traduire l’ensemble des ARN messagers présents dans la cellule et constitue donc l’usine de fabrication de l’ensemble des protéines nécessaires au bon fonctionnement cellulaire. Nous détaillerons dans cette section les acteurs et les principales étapes de la traduction eucaryote.
Les principaux acteurs de la traduction
Le processus de traduction est réalisé par l’action conjointe et coordonnée de plusieurs acteurs qui sont : • l’ARNm qui porte la séquence codant la protéine, • le ribosome qui valide le décodage et synthétise la protéine, • les ARN de transfert (ARNt) qui permettent de décoder chaque codon, • les facteurs protéiques qui vont permettre la réalisation des différentes étapes de la traduction.Avant de décrire les différents mécanismes impliqués dans la traduction, nous présenterons d’abord les 3 acteurs principaux : l’ARNm, le ribosome et les ARNt.
L’ARN messager
L’ARN messager mature qui sera traduit par le ribosome possède plusieurs éléments caractéristiques décrits sur la figure 1.2. C’est une molécule orientée qui se lit de l’extrémité 5’ vers l’extrémité A son extrémité 5’ on trouve un élément nommé la coiffe. La coiffe joue un rôle de protection de l’ARNm contre des exonucléases mais également un rôle essentiel de recrutement du ribosome lors de la première phase de la traduction : l’initiation [Furuichi 2001]. On trouve à la suite de la coiffe une région non codante (5’UTR pour untranslated region) qui ne sera donc pas traduite par le ribosome. Elle joue elle aussi un rôle dans l’initiation de la traduction. Cette séquence se termine par un codon initiateur, presque exclusivement AUG chez les eucaryotes, qui indique l’endroit où la traduction doit s’initier. A la suite de ces éléments on trouve une région codante de taille variable qui se termine par un codon particulier appelé codon stop. Il existe trois codons stop (UAA, UAG et UGA), mais ils donnent tous l’indication au ribosome d’interrompre le processus de traduction, de relâcher la protéine achevée et de se détacher de l’ARN messager. Enfin en aval du codon stop on trouve une seconde région non codante appelée 3’UTR puis un élément appelé queue polyA ou queue polyadénilée composée uniquement de bases adénines (environ 250 chez les mammifères, [Viphakone 2008]) qui joue un rôle lors de l’export de l’ARNm dans le cytoplasme ainsi qu’un rôle pour assurer sa stabilité dans le temps.
Le ribosome
Le ribosome est la machine moléculaire qui orchestre l’ensemble du processus de traduction. La structure détaillée (résolue à l’angström près grâce à la cristallographie X, cf. paragraphe 1.3) d’un ribosome eucaryote de levure n’a été élucidée qu’en 2010 par une équipe de l’Institut de génétique et de Biologie Moléculaire et Cellulaire de Starsbourg [Ben-Shem 2011], quelques années après le prix Nobel accordé pour la découverte de la structure du ribosome procaryote en 2000 [Ban 2000, Schluenzen 2000, Wimberly 2000].Le ribosome 1 est divisé deux sous-unités (SU), la grande avec un coefficient de sédimentation de 60 Svedberg (S) chez les eucaryotes et la petite sous-unité 40S. Les deux sous-unités sont elles même composées de protéines et d’ARN dit ribosomique (ARNr). L’ARN messager entre et ressort par un tunnel illustré en rouge sur la cartographie du ribosome de la figure 1.3, à droite 2 . Le ribosome 80S une fois assemblé sur l’ARN messager possède trois sites où vont se fixer les ARN de transfert porteurs des acides aminés pendant la traduction : les sites A, P et E.
Les ARN de transfert
Les ARNt sont de petites molécules d’ARN de 75 à 100 nucléotides. Ils servent d’adaptateurs entre l’ARNm et les acides aminés puisque ces molécules portent chacune un acide aminé. L’ARN de transfert est très structuré, sa structure secondaire forme une structure en L.La tige-boucle de l’anticodon va former la liaison codon-anticodon avec l’ARN messager lors de son passage dans les sites A,P et E du ribosome. Un anti-codon est un triplet de trois nucléotides complémentaires des nucléotides qui forment le codon de l’ARNm. Il se forme entre ces nucléotides des liaisons hydrogènes représentées par des traits pointillés sur la figure 1.4. A l’autre extrémité de l’ARNt, la tige-boucle acceptrice est liée à un acide aminé qui sera assemblé au reste de la chaine protéique pendant la traduction.
Etapes du processus
La traduction est un processus d’amplification où une molécule d’ARNm est à l’origine de la synthèse de plusieurs protéines. Il est important de noter que l’ARNm est traduit simultanément par plusieurs ribosomes à la suite les uns des autres, on parle alors de polysomes. Chaque ribosome accomplit un cycle de traduction qui se décompose en 4 étapes : • L’initiation durant laquelle les sous-unités ribosomiques s’assemblent sur l’ARNm et se positionnent sur le codon d’initiation. • l’élongation qui correspond à la lecture de l’ARNm et à la synthèse de la protéine. • la terminaison, étape à laquelle la protéine est relâchée. • la dissociation et le recyclage des sous-unités ribosomiques.
Initiation canonique de la traduction eucaryote
L’initiation canonique de la traduction eucaryote est une étape particulièrement complexe car elle implique un très grand nombre de facteurs d’initiation. Les différentes étapes de ce processus de traduction sont décrites sur la figure 1.5 3 . 1. L’initiation débute par l’activation de l’ARN messager via le recrutement du complexe d’initiation de la traduction eIF4F composé de trois sous-unités : eIF4A, eIF4E et eUF4G. Ces éléments interargissent directement avec le facteur protéique PAPB et la coiffe de l’ARNm pour former un premier complexe prêt à recruter le sous-unité 40S, nommé ARNm activé. 2. En parallèle se forme un complexe dit de pré-initiation (PIC) de coefficient de sédimentation 43S. On a d’abord formation d’un complexe ternaire comprenant l’ARNt initiateur ARNt-Met(i), le facteur d’initiation eucaryote eIF2 et une molécule de GTP (fig. 1.5(2.a)). Puis la petite sous-unité 40S s’associe avec les facteurs d’initiation eIF3, eIF1, eIF1A, eIF5 (2.b) et le complexe ternaire eIF2-ARNt-Met(i)-GTP pour aboutir au complexe 43S (2.c). 3. Le complexe de pré-initiation est recruté sur l’ARN messager activé. 4. Une fois ce complexe 48S formé, la sous-unité du ribosome peut alors balayer l’ARNm à la recherche du codon initiateur AUG. Les deux facteurs eIF1 et eIF1A sont importants dans ce mouvement de balayage. L’appariement des bases du codon initiateur AUG de l’ARNm et de l’anticodon de l’ARN de transfert initiateur ARNt-Met(i), lui-même positionné au site P du ribosome, permet d’aborder la phase finale de l’initiation. 5. Quand l’association codon start-anticodon est effectuée, l’hydrolyse du GTP lié à eIF2 permet l’éjection des facteurs d’initiation de la surface du complexe d’initiation 48S. La grande sous-unité ribosomique 60S peut alors s’associer par l’intermédiaire des facteurs eIF5 et eIF5B, permettant de former un ribosome 80S actif. Après une dernière étape de recyclage du facteur eIF2 sous la forme d’un complexe eIF2-GDP (qui sera recyclé pour s’associer dans un nouveau complexe ternaire), l’ensemble du système est prêt à entrer dans la phase d’élongation.