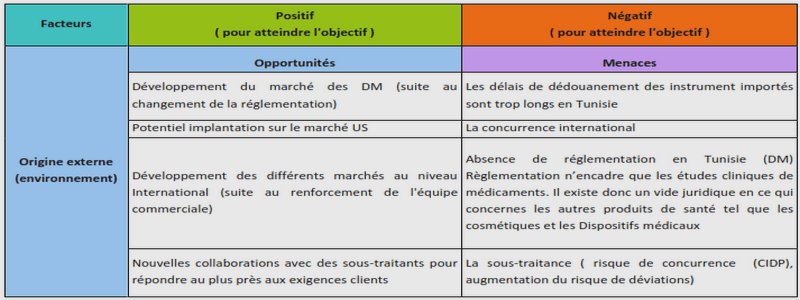
SENSIBILITE DE QUELQUES VARIETES DE BLE CULTIVEES AU RAVAGEUR Sitophilus zeamais
Motchulsky
Sitophilus zeamais Motchulsky
Systématique de l’insecte Régne : Animalia Phyllum : Arthropoda Hyperclasse : Hexapoda Classe : Insecta Sous classe : Ptérygote Ordre : Coléoptera Famille : Curculionidae Genre : Sitophilus Espèce : Sitophilus zeamais
Morphologie
Photo 1 : Adulte de Sitophilus zeamais (Source : Cissokho, 2010) Le genre Sitophilus est plus connu sous le nom de charançon. Les anciens l’appelaient calandres. Son corps a la forme d’un cylindre. Il se reconnait par la présence d’un rostre renflé triangulairement (Seck. 2009). Ce rostre porte des antennes en forme de massues à sa b ase. L’extrémité de ce rostre est équipée de pièces buccales de type broyeur. Chez la femelle le rostre est long et effilé tandis que chez le mâle, il est plus court et plus épais et moins lisse que celui de la femelle (Cissokho, 2010). Le corps, d’une longueur comprise entre 3 et 4mm, est couvert d’un tégument dur et rugueux. Les charançons subissent une métamorphose complète. Les larves sont des asticots charnus pourvus de pattes rudimentaires, de mandibules puissantes et d’yeux peu perfectionnés. Le prothorax de l’adulte est recouvert de piqure ronde Rostre Tête Thorax Tâches rougeâtres Abdomen 11 ou irrégulière. Il porte des ailes. Les ailes antérieures sont dures et tannées et rejoignent le long de la ligne centrale dorsale. Quant aux postérieures elles sont membraneuses quelquefois absentes. Les pattes portent à leurs extrémités des tarses à quatre (4) articles. Longtemps considéré comme une forme de S. oryzae de grande taille (larger strain) S. zeamais se distingue néanmoins d e ce d ernier. Certains auteurs (Kranz et al. 1977) ont proposé des critères d’ordre éthologique pour les différencier. S. oryzae (2 à 5 mm) est un bon voi lier, capable d’attaquer les céréales stockées depuis le champ. Il a une préférence pour les graines de petite taille comme le riz. Au contraire, S. zeamais (3 à 4 mm) vole très peu et préfère les graines de grande taille comme le maїs. D’autres clefs de distinction sont aussi utilisées. Adulte : de couleur brun-rouge foncé à noir, avec en général deux petites taches rougeâtres sur chaque élytre. Certains détails morphologiques permettent de distinguer S. zeamais de S. oryzae. En effet l’épisterne méta thoracique porte à ch acune de ces extrémités trois (3) rangées de ponctuations (deux seulement chez S. oryzae). La fossette frontale située entre les yeux est profonde et allongée. La ponctuation du pronotum ne laisse pas apparaitre de ligne médiane lisse, ou du moins celle-ci n’atteint pas la base de pronotum. Armature génitale : la face dorsale de l’édéage est creusée de deux sillons longitudinaux (bien visibles en lumière rasante), son extrémité est recourbée et pointue. Le huitième sternite de la femelle est en forme de Y aux branches convergentes et pointues à leur extrémité. Le dimorphisme sexuel est prononcé. La longueur du corps est en général supérieur à 3mm. Larve : de couleur blanche est semblable à celle de S. oryzae. Il ya quatre stades larvaires ; la nymphose s’effectue à l’intérieur de la graine. Lorsque plusieurs larves sont présentes dans le même grain, il n’en émerge qu’un seul adulte (cannibalisme). Ce v oilier est fréquemment rencontré dans les champs au moment de la maturation des grains. L’adulte est capable de parcourir plus de 800m pour atteindre la plante hôte. Le mâle est plus actif que la femelle. Il sécrète une phéromone d’agrégation identique à celle de S. oryzae : (4S,5R)-5-hydroxy-4-méthyl-3-heptanone (sitophinone), active envers les deux sexes Wakefield et al. (2005). I.1.3- Biologie 12 Sitophilus ne peut se reproduire que dans un grain dont les taux d’humidité est supérieur à 9,5%, sur une plage de températures allant de 15 à 35ºC. Les femelles adultes pondent environ 200 œufs à une vitesse de 20 à 30 par jour. Ces derniers sont déposés au fond de cavités creusées dans l’endosperme du gr ain et rebouchées à l’aide d’un tampon mucilagineux (Kossou and Bosque-Perez, 1998). A 18-20ºC, les œufs éclosent après 8 à 11 jours pour donner des larves de couleur blanche. Ces larves apodes, dures, légèrement courbées consomment l’endosperme du grain. Une seule larve se développe dans les grains de petite taille comme le blé. Cependant des graines de grosse taille comme le maїs supportent plusieurs spécimens. Les larves ne vivent jamais à l’air libre et se développent à l’intérieur du grain. Elles muent quatre (4) fois pour finalement se transformer en nymphes après 6 à 8 semaines. Les adultes émergent après 5 à 16 jours supplémentaires. Ils vivent environ 8 mois. S’ils sont dérangés ils feignent la mort en repliant leurs pattes sur leur corps. En effet les œufs ne sont guère affectés par les ovicides durant les six jours d’incubation. Le premier stade larvaire est vulnérable aux conditions de l’environnement (humidité relative, oxygène, densité de la population, température) (Longstaff, 1989 ; Hardman, 1978). Il peut y avoir jusqu’à 90% de mortalité à ce stade. Les survivants réussissent généralement à atteindre le stade adulte. Dans les conditions optimales de développement (27ºC et70% HR) les adultes vivent de 25 (femelles) à 30 semaines (mâles). Le cycle complet dure de 26 à 3 5 jours dans les zones tropicales.
Blé
Systématique du blé Embranchement : Spermaphytes Sous-embranchement : Angiospermes Super-classe : Monocotylédones Classe : Liliopsida Sous-classe : Commelinidae Groupe : Glumales 13 Ordre : Poales Famille : Poaceae Tribu : Triticeae Genre : Triticum L., 1753 Espèce : Triticum aestivum
Histoire et diversité des formes cultivées
Le blé fait partie des trois céréales dont les grains sont utilisés pour la nourriture humaine et animale. Cette m onocotylédone constitue la base alimentaire des populations du globe. L’origine du bl é (Triticum), du m aïs (Zea) et du riz (Oryza) semble être commune: étant donné les nombreux gènes communs deux à deux ou dans les trois genres. On pense que ces genres se sont diversifiés, il y a quelques 60 à 70 millions d’années (à la fin du secondaire) à partir d’une espèce ancestrale qui aurait contenu tous les gènes dispersés chez les trois espèces actuelles (Henry et Buyser, 2000). On retrouve donc sous le nom de blé des espèces variées du genre Triticum (Henry et Buyser, 2000). Les premiers indices d’une agriculture apparaissent vers 9000 ans avant Jésus-Christ dans le croissant fertile. On trouve dans les villages du dé but du Néolithique l’engrain (Triticum monococcum), l’amidonnier (Triticum dicoccum). Les formes sauvages identifiées de ces deux espèces seraient originaires du Proche-Orient et du Moyen-Orient. La céréaliculture se répand ensuite vers l’Europe, l’Asie et la vallée du Nil. Le froment est présent en Grèce il y a 6000 ans avant Jésus Christ et se propage par la méditerranée et le Danube. En Bretagne, on a trouvé des grains datant d’environ 5000 avant Jésus-Christ (Henry et Buyser, 2000). Les principaux caractères des espèces de blé que l’homme a cherché à sélectionner sont: la robustesse de l’axe de l’épi (qui ne doit pas se casser à la récolte), la séparation facile des enveloppes du grain, la grande taille des grains et la compacité des épis. Le blé constitue un groupe d’espèces polyploïdes (dont certaines espèces, polyploïdes, possèdent plusieurs jeux de chromosomes: 2n, 4n, 6n: avec n=7) (www.inra.fr/dpenv/doussd21.pdf). L’espèce actuelle diploïde est l’engrain (Triticum monococcum), rarement cultivé en France mais probablement domestiquée vers 8000 ans avant Jésus-Christ. 14 Elle est exclusivement issue de croisement de plusieurs blés sauvages diploïdes. Ces blés ont probablement donné naissance, vers 15000 ans avant Jésus-Christ, à un blé tétraploïde (4n) dont le représentant actuel est l’amidonnier (Triticum dicoccum) qui aurait été domestiqué vers 10000 ans avant Jésus-Christ. Cet amidonnier n’est pratiquement plus cultivé en France mais le reste dans les montagnes de l’Europe Centrale. Le blé dur actuel (Triticum durum), cultivé, est un blé tétraploïde issu d’une sélection réalisée à partir de l’amidonnier et qui daterait du temps de Jésus-Christ. Enfin, le blé tendre ou froment (Triticum æstivum) est hexaploïde (6n) et issu d’un double croisement récent entre une graminée sauvage diploïde et l’amidonnier tétraploïde. Aujourd’hui le premier critère dont il f aut tenir compte pour distinguer les espèces commerciales de blé est leur valeur d’utilisation industrielle. On distingue deux espèces de blé : le blé tendre et le blé dur. Ils se différencient par la friabilité de l’amande, qui est plus importante pour le blé tendre et permet la transformation en farine, alors que le blé dur est plus apte à se transformer en semoules. Le blé tendre est utilisé pour la panification, la pâtisserie, la biscuiterie, (Samuel et Modgil 1998). Il est panifiable. Le blé dur est utilisé pour les pâtes alimentaires, les semoules, les couscous.
Données botaniques
Sur le plant mature on note les racines fasciculées généralement superficielles. C’est une plante qui n’est pas capable d’aller chercher l’eau profondément dans le sol. Elle se développe rapidement et profite des pluies fréquentes. La tige creuse croît rapidement et donne des feuilles engainantes (qui entourent la tige par leur base), une à chaque nœud. Les feuilles sont simples, allongées et à nervures parallèles. L’axe de l’épi porte des épillets disposés alternativement d’un côté et de l’autre (alternes). L’épi est aplati pour les espèces cultivées. Un épillet comporte plusieurs fleurs protégées par 2 glumes. Chaque fleur, elle-même protégée par des glumelles, comporte 3 étamines et un ovaire avec 2 styles. Habituellement l’autofécondation est possible chez les blés. Ce sont les spermatozoïdes issus du pollen d’une fleur qui fécondent l’oosphère et la cellule centrale du sac embryonnaire de l’ovaire de cette même fleur. Les cellules sexuelles femelles sont protégées dans un sac embryonnaire fermé au sein d’un ovule, ce qui est un caractère d’Angiosperme (Henry et Buyser, 2000). Le fruit (grain de blé) est sec et indéhiscent et les enveloppes du fruit sont soudées à celles de la graine, ce qui fait que l’on nomme ce fruit-graine: caryopse. Les glumes et les glumelles sont perdues au battage. Les réserves sont contenues dans l’albumen composé à 70% d’amidon 15 et 15% de gluten. L’embryon n’a qu’un cotylédon et la plantule comporte un coléorhize, qui donnera la racine, et une coléoptile, qui donnera l’axe feuillet (Henry et Buyser, 2000). La germination du grain de blé dure entre 13 et 21 jours pendant lesquels le coléorhize sort de l’enveloppe du grain et donne une radicule d’où sont émises des racines primitives. Le coléoptile sort du grain et forme un étui protégeant les premières feuilles. L’axe portant le bourgeon terminal se développe en un rhizome (tige souterraine) dont la croissance s’arrête à 2 cm en-dessous de la surface du sol. Le développement des premières feuilles extérieures constitue la levée. Il apparaît un renflement dans la partie supérieure du rhizome qui grossit et forme le plateau de tallage. Le stade 3 feuilles est une phase repère pour le développement du blé. Des bourgeons se forment à l’aisselle des feuilles et donnent des pousses ou thalles. Chaque thalle primaire donne des thalles secondaires. A partir de la base du plateau de tallage apparaissent alors des racines secondaires ou adventives, qui seront à l’origine de l’augmentation du nombre d’épis (Henry et Buyser, 2000). Vient ensuite la montaison avec, au sommet du bourgeon Terminal, le début du développement de l’épi. Parallèlement, on assiste à l’allongement des entrenœuds. Le stade «épi à 1 cm» du plateau de tallage est caractérisé par une croissance active des thalles. Le plant de blé a besoin, durant cette phase, d’un important apport d’engrais azotés (Ichir, L.L et Ismaili, M. 2002). On assiste alors au «gonflement» qui correspond au développement de l’épi ou épiaison et à l’autofécondation. La floraison vient ensuite avec l’apparition des étamines en dehors des glumelles, le cycle s’achève par la maturation qui dure en moyenne 45 jours. Durant celle phase il y a migration des réserves depuis les parties vertes jusqu’aux grains, Quand le blé est mûr le végétal est sec et les graines des épis sont chargées de réserves (Henry et Buyser, 2000).
I.- GENERALITES |